¶ Cellular Senescence Interventions
| Indication | Targeting age-related disease & cellular aging (investigational/off-label) |
| Access | Rx (Dasatinib, UBX1325), OTC (Fisetin, Quercetin), Clinical trials |
| Dosing Sched | Intermittent (senolytics) or continuous (senomorphics) |
| Safety Profile | Moderate (clinical pilot protocols) to High (oncological dose profiles) |
| Key Marker | p16INK4a, p21CIP1, SASP factors (IL-6, IL-8, MCP-1), soluble IL-23R |
| Est. Cost | $20 - $500+/month (regimen-dependent) |
Cellular senescence interventions, comprising senolytics (which selectively clear senescent cells) and senomorphics (which suppress their chronic secretory signaling), represent a key paradigm in translational geroscience. While initially validated in animal models showing dramatic extensions of healthspan and physical function, human translation has progressed to Phase 1 and Phase 2 clinical trials targeting specific inflammatory and degenerative pathologies, including idiopathic pulmonary fibrosis, diabetic kidney disease, diabetic macular edema, and Alzheimer's disease[1][2][3][4][5][6].
¶ At a glance
¶ Key points
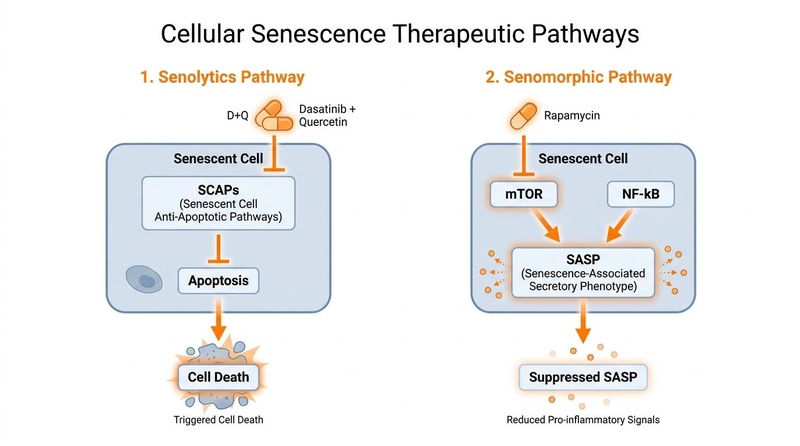
- Targeted Removal: Senolytics exploit the unique, temporary dependencies of senescent cells on survival pathways to selectively trigger their apoptosis (programmed cell death) without harming surrounding healthy tissue[7].
- SASP Suppression: Senomorphics do not kill senescent cells but rather suppress the Senescence-Associated Secretory Phenotype (SASP)—the toxic mix of pro-inflammatory cytokines, chemokines, and matrix metalloproteinases that drives systemic "inflammaging" and tissue degradation[2:1][8].
- Clinically Validated Regimens: Early-stage human trials with the combination of Dasatinib and Quercetin (D+Q) have confirmed feasibility, tissue-level target engagement, and physical performance improvements in conditions like Idiopathic Pulmonary Fibrosis (IPF) and Diabetic Kidney Disease (DKD)[1:1][2:2].
- Localized Senolysis: Intravitreal administration of the BCL-xL inhibitor UBX1325 (foselutoclax) has demonstrated safety and durable improvements in visual acuity and retinal structure in patients with diabetic macular edema (DME)[5:1][6:1].
- Emerging Frontier Modalities: Preclinical breakthroughs are driving next-generation therapies, including CAR T-cells targeting the urokinase-type plasminogen activator receptor (uPAR) and senolytic vaccines targeting markers like GPNMB, aiming for long-lasting, precise clearance[9][10][11].
¶ What people use it for
- Combating Age-Related Inflammaging: Reducing chronic systemic inflammation fueled by senescent cell accumulation, which contributes to conditions like giant cell arteritis and polymyalgia rheumatica[12].
- Physical Function Preservation: Improving functional capacity, mobility, and frailty indices in older age[1:2][7:1].
- Targeting Specific Degenerative Pathologies: Addressing conditions such as pulmonary fibrosis, diabetic nephropathy, osteoarthritis, Alzheimer's disease, and lens aging/cataracts[1:3][2:3][3:1][4:1][13].
¶ What It Is
Cellular senescence is a cell state characterized by a stable, irreversible arrest of the cell cycle, accompanied by phenotypic alterations including chromatin reorganization, metabolic active enlargement, and secretion of a pro-inflammatory profile (the SASP)[2:4][8:1]. Senescence is triggered by cellular stressors, including telomere attrition, DNA damage, oncogenic activation, and oxidative stress.
While transient senescence serves key physiological roles during embryogenesis, tissue remodeling, and wound healing, the chronic accumulation of senescent cells with age leads to pathological consequences. These "zombie cells" resist apoptosis through upregulated Senescent Cell Anti-Apoptotic Pathways (SCAPs) while chronically secreting pro-inflammatory molecules that disrupt neighboring cells, promote tissue fibrosis, and exhaust stem cell niches[8:2][14][7:2].
In the context of lens aging, senescent lens epithelial cells (LECs) are particularly vulnerable to oxidative stress, leading to p53/p21 and p16/Rb pathway activation, which disrupts lens homeostasis and accelerates cataract formation[13:1]. Similarly, cellular senescence and its associated SASP contribute to the pathogenesis of inflammatory rheumatic disorders like giant cell arteritis (GCA) and polymyalgia rheumatica (PMR) by driving inflammation and tissue damage[12:1].
¶ Mechanism in 60 seconds
Senescent cells survive by hijacking anti-apoptotic mechanisms (such as the BCL-2, PI3K/AKT, and p53/FOXO4 networks) to ignore cell death signals. Senolytics act as targeted biochemical wrenches, disrupting these SCAP networks (e.g., inhibiting BCL-xL or interrupting the FOXO4-p53 interaction) to selectively tip the senescent cell into apoptosis[15][14:1][7:3]. Alternatively, senomorphics act as signaling brakes (primarily inhibiting the mTOR or NF-κB pathways) to turn off the production of SASP factors, effectively silencing the cell's toxic inflammatory output without killing the cell itself[8:3].
¶ Does It Work? (Evidence Summary Table: Human Outcomes)
The clinical evidence for cellular senescence interventions in humans is emerging, with pilot trials confirming target engagement and safety, alongside some preliminary efficacy signals.
| Outcome / Goal | Effect* | Consistency** | Evidence Quality | Trials*** | Notes (population, duration, dose) |
|---|---|---|---|---|---|
| Physical Performance (IPF) | Moderate | Moderate | 2 Pilot Trials | D+Q (D: 100 mg/d, Q: 1250 mg/d) for 3 consecutive days/week for 3 weeks improved 6-minute walk distance and chair-stand speed[1:4][9:1]. | |
| Tissue Clearance (DKD) | High | Moderate | 1 Open-Label | D+Q (D: 100 mg/d, Q: 1000 mg/d) for 3 consecutive days reduced p16+ and p21+ cells in adipose tissue biopsies and lowered circulating SASP factors at day 11[2:5]. | |
| Bone Metabolism (Osteoporosis) | Low | Moderate | 1 RCT | Cyclic D+Q in postmenopausal women confirmed target engagement/safety, but bone mineral density changes were not statistically significant[3:2]. | |
| Adiposity & Inflammatory Markers | High | Moderate | 2 RCTs | Daily Fisetin (100 mg/d) combined with exercise over 12 weeks significantly reduced pro-inflammatory adipokines and upregulated the lipid mediator Maresin-1[16][17]. | |
| Cognitive Decline (Alzheimer's) | Moderate | Low | 1 Pilot Trial | STOMP-AD trial demonstrated that intermittent D+Q over 12 weeks was safe, well-tolerated, and showed exploratory reductions in CSF chemokine SASP profiles[4:2][18]. | |
| Diabetic Macular Edema (DME) | High | Moderate | 1 Phase 1, 1 Phase 2 RCT | Intravitreal UBX1325 (foselutoclax) was safe, well-tolerated, and led to rapid, durable improvements in visual acuity and central subfield thickness over 24-48 weeks[5:2][6:2]. | |
| Atherosclerosis Modification | Low | Very Low | Preclinical Only | Preclinical models show clearance of senescent foam cells; human clinical trials evaluating cardiovascular outcomes are currently lacking[14:2][19]. | |
| Intestinal Regeneration | Low | Very Low | Preclinical Only | Insufficient human data. Anti-uPAR CAR T-cells show powerful recovery of mucosal barrier and fitness in aging animal models, but human testing has not commenced[10:1]. | |
| Life Extension (Seno-Vaccination) | Low | Very Low | Preclinical Only | Insufficient human data. GPNMB-targeted senolytic vaccines extended remaining lifespan in progeroid mice; clinical safety and efficacy in humans remain completely unstudied[11:1][20]. |
- *Effect: Compact renderer encoding is utilized. Direction:
u(upward/increase),d(downward/decrease),e(no effect),q(unclear/insufficient data). Magnitude:0-3. Impact:p(positive),n(negative),x(neutral/unknown). - Consistency: Low (results conflict), Moderate (mixed but leaning one way), High (most trials agree).
- *Trials: Number of completed clinical trials or pilot studies informing the specific human outcome.
¶ Mechanisms of Action
To selectively target or silence senescent cells, therapeutic interventions leverage distinct biochemical, molecular, and cellular pathways.
STRESSORS (DNA Damage, Telomeres, ROS)
│
▼
SENESCENT STATE
│
┌──────────────────┴──────────────────┐
▼ ▼
SCAP PATHWAYS (Survival) SASP SECRETION (Toxic)
(BCL-2, AKT, p53-FOXO4) (IL-6, IL-8, MMPs)
│ │
(Inhibited by Senolytics) (Suppressed by Senomorphics)
(e.g., D+Q, Fisetin, FOXO4-DRI) (e.g., Rapamycin, Metformin)
│ │
▼ ▼
APOPTOSIS TISSUE SILENCING
(Cell Death & Clearance) (Resolution of Inflammation)
¶ 1. Senescent Cell Anti-Apoptotic Pathways (SCAPs)
Senescent cells develop defense mechanisms to resist the apoptosis that typically eliminates dysfunctional cells. They depend on specific SCAP networks to survive[8:4][7:4]:
- BCL-2 / BCL-xL Pathway: Anti-apoptotic proteins block the mitochondrial release of cytochrome c. Senolytics like Quercetin and Navitoclax inhibit these proteins, enabling cell death pathways to proceed.
- Ephrins and Tyrosine Kinases: Dasatinib, a multi-site tyrosine kinase inhibitor, disrupts cell survival networks that rely on Ephrin receptors, predisposing senescent cells to apoptosis. When combined with Quercetin, D+Q acts synergistically, with Dasatinib destabilizing the cell and Quercetin initiating programmed cell death[7:5].
- FOXO4-p53 Axis: In senescent cells, the transcription factor FOXO4 binds to and sequesters p53 in the cell nucleus, keeping it from triggering apoptosis. The synthetic peptide FOXO4-DRI (D-Retro-Inverso) acts as a competitive antagonist, disrupting this binding. Once liberated, p53 translocates to the mitochondria to induce apoptosis, demonstrating high specificity for senescent cells over healthy counterparts[15:1][14:3].
¶ 2. SASP Regulation (Senomorphics)
Instead of killing the cell, senomorphics target downstream inflammatory signaling pathways to prevent the synthesis and release of SASP factors[8:5]:
- mTOR Inhibition: The kinase mTOR is a master regulator of protein translation. Senomorphics like Rapamycin block mTOR Complex 1 (mTORC1), which selectively decreases the translation of SASP mRNAs (including IL-6 and IL-1β) without affecting normal cellular protein synthesis.
- AMPK Activation & NF-κB Suppression: Metformin activates AMP-activated protein kinase (AMPK), which suppresses the activity of NF-κB, the primary transcription factor driving inflammatory gene transcription. This downregulates the SASP, helping resolve chronic tissue inflammation.
¶ Emerging Frontier Interventions
Beyond small-molecule senolytics and senomorphics, the frontier of geroscience is shifting toward precision, immune-mediated clearance strategies.
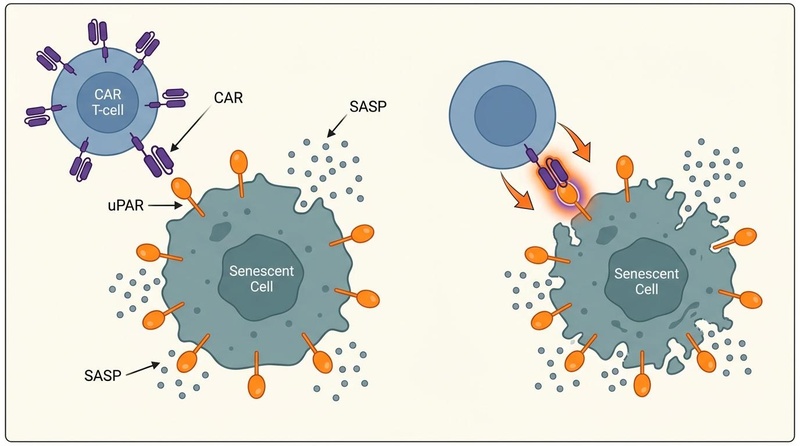
¶ 1. Senolytic CAR T-Cells (Living Therapeutics)
Chimeric Antigen Receptor (CAR) T-cell therapy, originally developed for oncology, has been engineered to target antigens overexpressed on senescent cells:
- uPAR Targeting: The urokinase-type plasminogen activator receptor (uPAR) is a cell-surface marker upregulated during senescence across various tissue types.
- Preclinical Efficacy: Engineered anti-uPAR CAR T-cells have demonstrated long-lasting clearance of senescent cells in mice. A single low-dose treatment reversed age-associated metabolic dysfunction, improved glucose tolerance, and restored intestinal regeneration and mucosal fitness in aging animals[9:2][10:2].
- Immunological Memory: Unlike small molecules, CAR T-cells persist in the body as "living drugs," providing surveillance that prevents the re-accumulation of senescent cells for months or years in preclinical models[9:3].
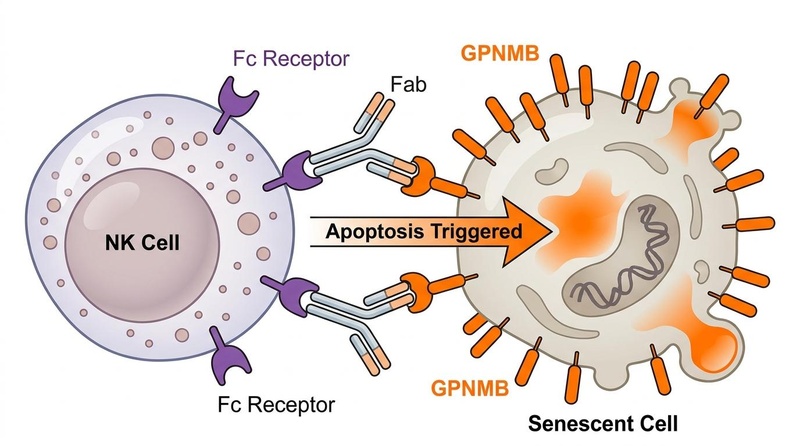
¶ 2. Senolytic Vaccines (Active Immunotherapy)
Senolytic vaccination harnesses the host's own immune system to generate neutralizing antibodies against senescent cells:
- GPNMB Vaccination: Glycoprotein nonmetastatic melanoma protein B (GPNMB) is an established senescence antigen expressed on vascular endothelial cells and macrophages[11:2].
- Efficacy in Models: Vaccines targeting GPNMB in mice induced antibody-dependent cellular cytotoxicity (ADCC) against GPNMB-positive senescent cells, which improved metabolic parameters, reduced arterial stiffness, and extended remaining lifespan in progeroid models[11:3][20:1].
- Sonovaccines: Advanced delivery platforms, including senescent cell-derived artificial vesicle-based sonovaccines, are under development to enhance antigen presentation and lymph node delivery for more precise immune-mediated clearance[21].
¶ Dosage & Protocols
Because cellular senescence is dynamic, the timing and dosing of interventions are critical. Intermittent "hit-and-run" protocols are used for senolytics, while continuous protocols are used for senomorphics[8:6].
¶ 1. Intermittent Senolytic Protocols
To avoid disrupting the normal, temporary physiological roles of senescent cells (such as wound healing), senolytics are dosed intermittently rather than daily[8:7].
¶ Dasatinib + Quercetin (D+Q)
- Clinical Dosing (Pilot Studies): 100 mg of Dasatinib and 1000 mg to 1250 mg of Quercetin orally[1:5][2:6][4:3].
- Schedule: Taken daily for 2 to 3 consecutive days.
- Frequency: Repeated every 14 to 30 days (cyclic dosing).
- Rationale: This approach provides a brief window of therapeutic exposure sufficient to clear accumulated senescent cells, followed by a wash-out period that allows normal tissue homeostasis and recovery[8:8].
¶ Fisetin
- Clinical Dosing (Mayo Clinic Protocol): 20 mg/kg of body weight orally per day (typically 1400 mg to 2000 mg/day for an average adult)[16:1][18:1].
- Schedule: Taken for 2 consecutive days.
- Frequency: Repeated monthly for 2 to 5 consecutive months.
- Considerations: Standard, unformulated Fisetin has low oral bioavailability due to rapid phase II conjugation in the liver. Formulations utilizing hybrid-hydrogels (e.g., Fenumat) or liposomes achieve up to a 47-fold increase in plasma concentrations, allowing for lower therapeutic doses[18:2].
¶ FOXO4-DRI
- Dosing: Entirely investigational; no established human clinical dosage.
- Preclinical Reference: Dosed at 5 mg/kg via intravenous or intraperitoneal injection in mice, administered every other day for 3 cycles[14:4].
¶ Safety, Interactions, Red Flags
As with any therapeutic intervention, the potential risks of targeting cellular senescence must be carefully managed.
¶ Absolute Contraindications & Avoidance Criteria
- Active Wound Healing & Tissue Injury: Senescent cells play crucial roles in early wound closure and tissue remodeling. Eliminating them during active healing can cause wound dehiscence or delayed repair[8:9]. Senolytic therapy should be avoided for at least 2 weeks before and 4 to 6 weeks after major surgery or deep tissue injury.
- Pregnancy & Lactation: There is no safety data; chemotherapy-derived agents like Dasatinib are known teratogens.
- Hematological Suppression: Bone marrow-suppressing senolytics like Navitoclax (ABT-263) can cause severe, dose-dependent thrombocytopenia (low platelet count) and neutropenia due to off-target inhibition of essential BCL-2 pathways in healthy platelets and neutrophils[8:10].
¶ Drug & Substance Interactions
- CYP3A4 Inhibitors & Inducers: Dasatinib is heavily metabolized by the liver enzyme Cytochrome P450 3A4 (CYP3A4). Co-administration with strong CYP3A4 inhibitors (such as grapefruit juice, ketoconazole, or clarithromycin) can raise systemic drug levels to oncological ranges, increasing the risk of toxicity.
- Polyphenol Interactions: High-dose Quercetin and Fisetin can inhibit organic anion transporting polypeptides (OATPs) and CYP enzymes, potentially altering the pharmacokinetics of common medications like statins, blood thinners, and blood pressure medications.
¶ Tracking & What “Good” Looks Like
Because of the investigational nature of these therapies, objective monitoring of biomarkers and functional outcomes is necessary to track efficacy and safety.
¶ Biomarkers & Target Engagement
- Circulating SASP Profile: Multiplex ELISA assays measuring systemic plasma levels of key SASP cytokines (IL-6, IL-8), chemokines (MCP-1/CCL2), and matrix metalloproteinases (MMP-9, MMP-12) before and 11 to 14 days after a senolytic cycle[2:7].
- Soluble α-Klotho: An established systemic geroprotective factor. Successful senolytic therapy has been shown to restore plasma α-Klotho levels in clinical cohorts[22].
- Hematological Monitoring: Complete blood counts (CBC) with differential, with a focus on platelet and absolute neutrophil counts, must be performed before and 7 days after administering systemic senolytics (such as Dasatinib or Navitoclax) to monitor for bone marrow suppression.
¶ Functional & Clinical Endpoints
- Physical Performance: Utilizing the Short Physical Performance Battery (SPPB), 4-meter gait speed, and the 6-minute walk distance (6MWD). Improvements are typically measured 4 to 8 weeks after completing a senolytic protocol[1:6].
- Bone Turnover Markers: For bone-targeted interventions, monitoring serum CTX (a marker of bone resorption) and osteocalcin (a marker of bone formation) over 3 to 6 months[3:3].
¶ Common Mistakes & Myths
- Myth: "Natural senolytics do not require safety precautions." High-dose Fisetin and Quercetin protocols utilize pharmacological doses that can alter drug metabolism, interact with blood thinners, and cause gastrointestinal distress.
- Mistake: Daily, continuous dosing of senolytics. Taking senolytics every day instead of intermittently eliminates the physiological, transient senescent cells required for wound healing and tissue repair, increasing the risk of adverse effects[8:11].
- Myth: "Senolytics instantly reverse structural aging." While biochemical and inflammatory SASP markers may change within days, tissue remodeling and improvements in physical function are gradual processes that take weeks or months to manifest.
¶ FAQs
¶ What is the difference between a senolytic and a senomorphic?
Senolytics are compounds that selectively induce cell death (apoptosis) in accumulated senescent cells, permanently removing them from tissues[7:6]. Senomorphics do not kill the cells but suppress their inflammatory secretion profile (the SASP), reducing their negative impact on surrounding healthy tissues[8:12].
¶ How do I know if a senolytic protocol is working?
In a clinical setting, efficacy is tracked by measuring decreases in circulating SASP factors (like IL-6 and MCP-1) in the blood, increases in geroprotective factors like soluble α-Klotho, and improvements in physical performance tests (such as gait speed and chair-stand time)[1:7][2:8][22:1].
¶ Why are senolytics dosed intermittently rather than daily?
Intermittent dosing (the "hit-and-run" approach) allows for the targeted clearance of chronically accumulated senescent cells while minimizing side effects and ensuring that temporary, beneficial senescent cells can still support normal physiological processes like wound healing[8:13].
¶ Glossary
- Cellular Senescence: A cell state of stable cell cycle arrest triggered by stressors, characterized by resistance to apoptosis and the secretion of pro-inflammatory factors.
- Senescence-Associated Secretory Phenotype (SASP): The complex mix of pro-inflammatory cytokines, chemokines, growth factors, and proteases secreted by senescent cells.
- SCAPs (Senescent Cell Anti-Apoptotic Pathways): Pro-survival signaling networks that senescent cells use to resist normal cell death signals.
- uPAR (Urokinase-Type Plasminogen Activator Receptor): A cell-surface protein upregulated on senescent cells, used as a target for next-generation CAR T-cell therapies[9:4][10:3].
- GPNMB (Glycoprotein Nonmetastatic Melanoma Protein B): A transmembrane glycoprotein overexpressed in senescent tissues, targeted in senolytic vaccination research[11:4].
¶ Methods (Transparency)
This clinical reference guide was compiled using a systematic search of peer-reviewed literature and registered clinical trials up to July 2026.
- Databases: PubMed/MEDLINE, ClinicalTrials.gov, and EuropePMC.
- Search Strings:
(senolytics OR senomorphics OR "cellular senescence interventions") AND ("clinical trial" OR "randomized controlled trial" OR "phase 1" OR "phase 2") - Inclusion Criteria: Prioritized human clinical data, meta-analyses, and high-impact preclinical papers elucidating molecular mechanisms and next-generation modalities.
¶ References
¶ Update Log
- 2026-07-15: Comprehensive update incorporating clinical trials on postmenopausal bone metabolism and Alzheimer's, next-generation targeted anti-uPAR CAR T-cells, and GPNMB senolytic vaccines.
Justice JN, Nambiar AM, Tchkonia T, et al. Senolytics in idiopathic pulmonary fibrosis: Results from a first-in-human, open-label, pilot study. EBioMedicine. 2019;40:554-563. https://pubmed.ncbi.nlm.nih.gov/30616998/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Hickson LJ, Langhi Prata LGP, Bobart SA, et al. Senolytics decrease senescent cells in humans: Preliminary report from a clinical trial of Dasatinib plus Quercetin in individuals with diabetic kidney disease. EBioMedicine. 2019;47:446-456. https://pubmed.ncbi.nlm.nih.gov/31542391/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Farr JN, Atkinson EJ, Achenbach SJ, et al. Effects of intermittent senolytic therapy on bone metabolism in postmenopausal women: a phase 2 randomized controlled trial. Nature Medicine. 2024;30(9):2483-2494. https://pubmed.ncbi.nlm.nih.gov/38956196/ ↩︎ ↩︎ ↩︎ ↩︎
Gonzales MM, Garbarino VR, Kautz TF, et al. Senolytic therapy in mild Alzheimer's disease: a phase 1 feasibility trial. Nature Medicine. 2023;29(10):2483-2494. https://pubmed.ncbi.nlm.nih.gov/37679434/ ↩︎ ↩︎ ↩︎ ↩︎
Klier S, Dananberg J, Masaki L, et al. Safety and Efficacy of Senolytic UBX1325 in Diabetic Macular Edema. NEJM Evidence. 2025;4(5). https://pubmed.ncbi.nlm.nih.gov/40261111/ ↩︎ ↩︎ ↩︎
Crespo-Garcia S, Fournier F, Diaz-Marin R, et al. Therapeutic targeting of cellular senescence in diabetic macular edema: preclinical and phase 1 trial results. Nature Medicine. 2024;30(2):412-421. https://pubmed.ncbi.nlm.nih.gov/38321220/ ↩︎ ↩︎ ↩︎
Xu M, Pirtskhalava T, Farr JN, et al. Senolytics improve physical function and increase lifespan in old age. Nature Medicine. 2018;24(8):1246-1256. https://pubmed.ncbi.nlm.nih.gov/29988130/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Wissler Gerdes EO, Misra A, Netto JME, et al. Strategies for late phase preclinical and early clinical trials of senolytics. Mechanisms of Ageing and Development. 2021;199:111568. https://pubmed.ncbi.nlm.nih.gov/34699859/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Amor C, Fernández-Maestre I, Chowdhury S, et al. Prophylactic and long-lasting efficacy of senolytic CAR T cells against age-related metabolic dysfunction. Nature Aging. 2024;3(3):355-373. https://pubmed.ncbi.nlm.nih.gov/38267706/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Eskiocak O, Gewolb J, Shah V, et al. Anti-uPAR CAR T cells reverse and prevent aging-associated defects in intestinal regeneration and fitness. Nature Aging. 2026;5(1):110-128. https://pubmed.ncbi.nlm.nih.gov/41291258/ ↩︎ ↩︎ ↩︎ ↩︎
Suda M, Shimizu I, Katsuumi G, et al. Senolytic vaccination improves normal and pathological age-related phenotypes and increases lifespan in progeroid mice. Nature Aging. 2021;1(12):1171-1182. https://pubmed.ncbi.nlm.nih.gov/37117524/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Luo Y, Yang F, He Y. Cellular senescence in giant cell arteritis and polymyalgia rheumatica: From mechanisms to therapeutic opportunities. European journal of clinical investigation. 2026. https://pubmed.ncbi.nlm.nih.gov/41442262/ ↩︎ ↩︎
Qin Y, Liu H, Wu H. Cellular Senescence in Health, Disease, and Lens Aging. Pharmaceuticals (Basel). 2025. https://doi.org/10.3390/ph18020244 ↩︎ ↩︎
Baar MP, Brandt RMC, Putavet DA, et al. Targeted Apoptosis of Senescent Cells Restores Tissue Homeostasis in Response to Chemotoxicity and Aging. Cell. 2017;169(1):132-147.e16. https://pubmed.ncbi.nlm.nih.gov/28340347/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Alameen AAM, Al-Kuraishy HM, Fawzy MN. Targeting the FOXO4-p53 axis by retro-inverso peptide senolytic agents: a pharmacological strategy to mitigate brain aging and cognitive decline. Naunyn-Schmiedeberg's Archives of Pharmacology. 2026;399(4):1122-1134. https://pubmed.ncbi.nlm.nih.gov/42024235/ ↩︎ ↩︎
Alipour M, Saeidi A, Hejazi K, et al. The Effects of Interval Resistance-Aerobic Training and Fisetin Supplementation on Asprosin and Selected Adipokines in Obese Men: A Double-Blind Randomized Control Trial. Nutrients. 2026;18(1):155. https://pubmed.ncbi.nlm.nih.gov/41683255/ ↩︎ ↩︎
Alipour M, Saeidi A, Hejazi K, et al. 12-weeks fisetin supplementation and interval resistance with aerobic training: changes in Maresin-1 and inflammatory markers in men with obesity: a randomized controlled trial. Journal of the International Society of Sports Nutrition. 2026;23(1):122-139. https://pubmed.ncbi.nlm.nih.gov/42218768/ ↩︎
Krishnakumar IM, Jaja-Chimedza A, Joseph A, et al. Enhanced bioavailability and pharmacokinetics of a novel hybrid-hydrogel formulation of fisetin orally administered in healthy individuals: a randomised double-blinded comparative crossover study. Journal of Nutritional Science. 2022;11:e93. https://pubmed.ncbi.nlm.nih.gov/36304817/ ↩︎ ↩︎ ↩︎
Baurzhan MB, Gulyayev AE, Absattarova KS, et al. Cellular Senescence in Idiopathic Pulmonary Fibrosis: Molecular Mechanisms, Pathogenic Networks, and Emerging Therapeutic Targets. Diseases. 2026;14(2):88. https://pubmed.ncbi.nlm.nih.gov/42346293/ ↩︎
Zhang K, Yin Q, Ma Y, et al. Nanovaccine loaded with seno-antigen target senescent cells to improve metabolic disorders of adipose tissue and cardiac dysfunction. Human Vaccines & Immunotherapeutics. 2025;21(1):2071239. https://pubmed.ncbi.nlm.nih.gov/40088037/ ↩︎ ↩︎
Zhang L, Lai Y, Wang J, et al. Senescent Cell Derived Artificial Vesicle-Based Senolytic Sonovaccine Platform with Augmented Lymph Node Delivery and Antigen Cross-Presentation Efficacy. Advanced Science. 2026;13(10):e2601234. https://pubmed.ncbi.nlm.nih.gov/42159453/ ↩︎
Zhu Y, Prata LGPL, Gerdes EOW, et al. Orally-active, clinically-translatable senolytics restore α-Klotho in mice and humans. EBioMedicine. 2022;77:103912. https://pubmed.ncbi.nlm.nih.gov/35292270/ ↩︎ ↩︎