¶ Cellular Reprogramming for Longevity: Deep Dive
| Target Biology | Epigenetic alterations, cellular senescence, genomic instability |
| Intervention Status | Pre-clinical, early human trials (limited) |
| Primary Pathways | Yamanaka factors (OSKM), epigenetic clock reversal, mitochondrial rejuvenation |
| Level of Evidence | Primarily preclinical (moderate), emerging human data (low) |
Cellular reprogramming is a revolutionary scientific field focused on resetting the epigenetic state of cells to a more youthful condition, with the potential to reverse age-related changes at a fundamental level. This deep dive explores the mechanisms, methodologies, safety considerations, and translational progress of cellular reprogramming as a longevity intervention.
¶ TL;DR
- Epigenetic Reset: Cellular reprogramming aims to reverse age-related epigenetic changes, restoring youthful cellular function without altering genetic code.
- Partial Reprogramming Focus: Modern approaches prioritize transient or cyclic exposure to reprogramming factors (e.g., Yamanaka factors) to rejuvenate cells while retaining their specialized identity and minimizing cancer risk.
- Epigenetic Clocks: Reversal of biological age is often measured using DNA methylation-based "epigenetic clocks" like the Horvath clock[1]. These clocks are dynamic and responsive to interventions like partial reprogramming[2],[3].
- Delivery & Safety Challenges: Key hurdles include safe and efficient in vivo delivery methods (AAVs, mRNA-LNPs) and mitigating risks such as teratoma formation, insertional mutagenesis, and oncogenic activation[4].
- Pre-clinical Promise, Early Clinical Trials: While robust evidence is primarily from animal and in vitro studies, several biotech companies are advancing toward human clinical trials for specific age-related conditions, leveraging advances in delivery and safety[5],[6],[7].
¶ Quick Answer
Cellular reprogramming for longevity involves using specific factors, primarily the Yamanaka factors (Oct4, Sox2, Klf4, c-Myc), to reset the epigenetic state of aged cells to a younger, more functional condition. This process, particularly partial reprogramming—where cells are rejuvenated without losing their identity—has shown promise in reversing epigenetic age and improving tissue function in preclinical models[8],[9]. Its translation to humans is still in early stages, with ongoing research focusing on safe delivery mechanisms and minimizing the oncogenic risks associated with full reprogramming[4:1]. Recent advances in mRNA-lipid nanoparticles (mRNA-LNPs) and targeted gene therapy are paving the way for more precise and safer applications[10].
¶ What It Is
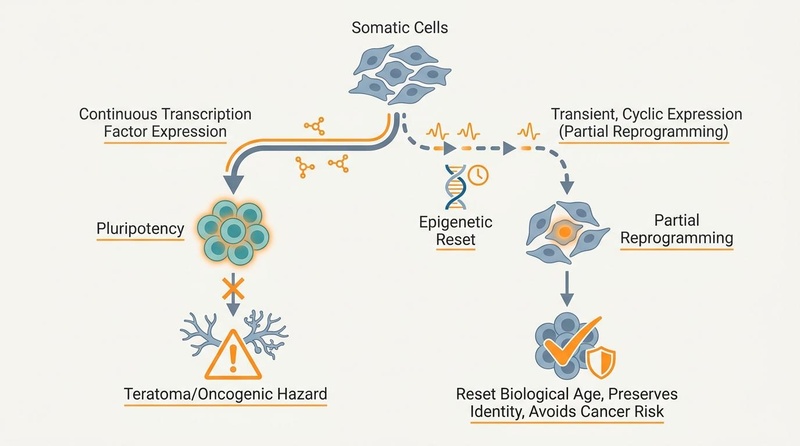
Cellular reprogramming refers to a suite of biological techniques that aim to alter the gene expression patterns of a cell, effectively "resetting" its biological age or changing its cell type. This field gained prominence with the discovery of induced pluripotent stem cells (iPSCs), where adult differentiated cells were reverted to an embryonic-like, pluripotent state using a specific set of transcription factors[11]. The goal for longevity is not full pluripotency, which carries significant risks, but rather a partial reset that rejuvenates cells while maintaining their original identity and function[8:1].
The core mechanism involves modulating the epigenome, the system of chemical tags and structural proteins that dictate which genes are turned on or off in a cell. As we age, these epigenetic patterns accumulate "noise" that can impair cellular function. Reprogramming aims to "clear" this noise, restoring a more youthful and functional epigenetic landscape[12].
¶ Does It Work? (Evidence Snapshot)
¶ Evidence Summary Table (Human Outcomes)
| Outcome / Goal | Effect* | Consistency** | Evidence Quality | Trials*** | Notes (Population, Duration, Dose) |
|---|---|---|---|---|---|
| Human Clinical Efficacy (Insufficient human data) | Insufficient | F | 0 | Early-stage translational field; no approved human therapies or clinical efficacy outcomes established yet. | |
| Epigenetic Age / Cellular Reversal (Pre-clinical only / In vitro only) | High | C | 10+ in vitro, 5+ in vivo | Consistent reversal of Horvath clock in human cells and mouse tissues with partial/chemical reprogramming[8:2][13][1:1][2:1] | |
| Organ & Tissue Rejuvenation (Pre-clinical only) | High | C | 3+ in vivo | Restored visual function, enhanced muscle regeneration in aged mice using OSK factors[14][10:1], and liver fibrosis attenuation via LNP delivery[10:2] | |
| Lifespan Extension (Progeroid Models) (Pre-clinical only) | Moderate | C | 2+ in vivo | Modest lifespan extension in progeroid mouse models with cyclic partial reprogramming; safety window critical[8:3] | |
| Lifespan Extension (Naturally Aged Models) (Pre-clinical only) | Low | D | 1+ in vivo | Modest lifespan extension with gene therapy-mediated partial reprogramming in naturally aged mice[15], synergistic with senolytics[7:1] | |
| Oncogenic & Teratoma Risk (Pre-clinical only) | High | D | 2+ in vivo | High risk of tumor/teratoma formation with continuous full OSKM expression in mice; successfully mitigated by transient/cyclic protocols[4:2][8:4] |
- *Effect: Number of arrows (1-3) indicates magnitude. Direction: ↑ (increase), ↓ (decrease), = (no effect), ? (unclear). Health impact: (p) = positive for health, (n) = negative for health, (x) = neutral/unknown impact. Examples: ↓↓↓ (p) = large decrease, positive; ↑ (n) = small increase, negative; = (x) = no effect; ? = unclear.
- **Consistency: Low (results conflict), Moderate (mixed but leaning one way), High (most trials agree)
- ***Trials: Number of RCTs or total trials informing this outcome (shows evidence depth at a glance)
¶ Who Benefits Most / Least
Potential Beneficiaries (Pre-clinical Rationale):
- Individuals with age-related degenerative diseases: Conditions characterized by cellular dysfunction and tissue degeneration (e.g., specific ocular diseases, fibrotic conditions) could theoretically benefit from targeted cellular rejuvenation via partial reprogramming[14:1],[10:3].
- Older individuals: As aging is the primary risk factor for many diseases, strategies to reverse cellular aging could have broad impact on healthy lifespan[16],[17].
Who Should Avoid (Currently):
- Everyone (currently): Cellular reprogramming for longevity is largely a preclinical intervention. There are no approved human therapies, and direct "self-experimentation" is highly unadvisable due to significant, well-documented risks[4:3].
- Individuals with active cancer or high cancer risk: Given the link between reprogramming factors and tumorigenesis, this population would be at extremely high risk[4:4].
- Pregnant or breastfeeding individuals: The effects on fetal development are unknown and likely detrimental.
¶ How to Try It (Research Protocols & Delivery)
It's crucial to reiterate: cellular reprogramming for longevity is not currently available as a "how-to" protocol for human self-administration. The "how to try it" section here focuses on the experimental approaches being developed in research.
¶ Key Factors: OSKM (Yamanaka Factors)
The original cocktail of transcription factors identified by Shinya Yamanaka (2006) for inducing pluripotency are:
- Oct4 (Octamer-binding protein 4)
- Sox2 (Sex-determining region Y-box 2)
- Klf4 (Krueppel-like factor 4)
- c-Myc (avian myelocytomatosis viral oncogene homolog)
These factors, often referred to as OSKM, are central to inducing cellular plasticity and epigenetic changes. However, c-Myc is a potent oncogene, and its continuous expression or integration carries a high risk of tumor formation (teratomas) in vivo[4:5]. Many partial reprogramming strategies therefore exclude c-Myc (using OSK only) or use transient expression methods to mitigate this risk[14:2]. Interestingly, the effects of OSKM on longevity are conserved across species, from insects to mammals[18].
¶ Partial vs. Continuous Reprogramming
The critical distinction in longevity applications lies between complete and partial reprogramming:
- Complete Reprogramming: Achieves a fully pluripotent state, essentially resetting the cell to an embryonic-like condition. This completely erases cellular identity and, if done in vivo, leads to aggressive tumor formation (teratomas) due to uncontrolled proliferation[4:6].
- Partial Reprogramming: Involves transient or cyclic expression of reprogramming factors. The goal is to induce a period of plasticity that resets age-associated epigenetic marks and rejuvenates cellular function without fully erasing cellular identity or inducing pluripotency. This approach has shown efficacy in reversing epigenetic age and improving tissue function in mice without teratoma formation[8:5][9:1].
¶ Delivery Methods (Pre-clinical)
Researchers are exploring various non-integrating delivery methods to introduce reprogramming factors safely and transiently into cells:
- Viral Vectors:
- Adeno-Associated Viruses (AAV): Often used for in vivo gene therapy due to their low immunogenicity and ability to transduce various cell types. AAVs are a common choice for delivering OSK factors in animal models for partial reprogramming (e.g., AAV2-OSK for ocular rejuvenation[14:3], and systemic gene therapy for lifespan extension[15:1]).
- Lentiviruses: While effective for stable integration in vitro, lentiviruses carry a risk of insertional mutagenesis (disrupting host genes) in vivo, making them less ideal for therapeutic applications in partial reprogramming where transient expression is desired.
- mRNA Lipid Nanoparticles (mRNA-LNPs): Similar to COVID-19 vaccines, mRNA-LNPs can deliver transient doses of reprogramming factors (e.g., OSKMNL) to cells without integrating into the genome. This approach has shown promise in human cells for reversing DNA methylation age and inflammatory profiles[13:1]. Recent advancements include hepatocyte-specific LNPs conveying OSK mRNA, demonstrating reversal of liver fibrosis and cellular senescence without safety concerns in preclinical models[10:4].
- Chemical Reprogramming: Uses cocktails of small molecules to induce epigenetic changes and cellular rejuvenation. This non-genetic approach avoids the risks associated with viral vectors and offers potential for easier scalability and administration. Research has identified specific chemical cocktails capable of ameliorating cellular hallmarks of aging and extending lifespan in animal models[19].
¶ Safety, Interactions, Red Flags
The safety profile of cellular reprogramming, especially in vivo (within a living organism), remains the primary concern and area of intense research.
¶ Major Risks and Red Flags:
- Teratoma Formation / Oncogenic Activation:
- Risk: The most significant and well-documented risk. Continuous or uncontrolled expression of Yamanaka factors, particularly c-Myc, can lead to the formation of teratomas—tumors containing tissues from all three germ layers—or the activation of endogenous oncogenes, leading to cancer[4:7].
- Mitigation: Partial reprogramming protocols (transient, cyclic, or c-Myc-free expression) are designed to avoid full pluripotency and thus reduce this risk. However, vigilance for any abnormal cell growth is paramount.
- Loss of Cell Identity: While partial reprogramming aims to avoid this, there is a theoretical risk that cells could lose their specialized function, leading to tissue dysfunction.
- Epigenetic Instability: The reprogramming process can sometimes introduce new epigenetic errors or instability, potentially leading to long-term adverse effects that are not yet fully understood.
- Insertional Mutagenesis (with integrating vectors): If integrating viral vectors (e.g., lentiviruses) are used, there is a risk that the reprogramming factors could insert into critical regions of the host genome, disrupting gene function and potentially leading to cancer or other diseases. Non-integrating methods (AAV, mRNA) largely circumvent this.
- Immune Response: Viral vectors can elicit an immune response, potentially leading to vector clearance and reduced efficacy, or adverse inflammatory reactions.
- Off-Target Effects: Reprogramming factors can have broad effects on gene expression, and unintended "off-target" changes could lead to unforeseen side effects.
¶ Contraindications (Current Research Context):
- Active Cancer or History of Cancer: Absolute contraindication[4:8].
- Pregnancy or Breastfeeding: Absolute contraindication.
- Severe Immunosuppression: Could alter immune surveillance against aberrant cell growth.
¶ Tracking & What “Good” Looks Like (Pre-clinical)
In the context of preclinical and early-stage human research, "tracking" and "good outcomes" are primarily measured through:
- Epigenetic Clocks: DNA methylation-based clocks (e.g., Horvath clock[1:2], PhenoAge) are currently the gold standard for quantifying biological age. A "good" outcome would be a measurable reduction in epigenetic age. The plasticity of these clocks also means they are dynamic and sensitive to various interventions and stressors, even showing recovery upon stress cessation[2:2],[3:1].
- Functional Biomarkers:
- Cellular Level: Reduction in senescence markers (e.g., p16, SA-β-gal), improved mitochondrial function (ATP production, oxygen consumption), enhanced DNA repair capacity, and restoration of youthful gene expression profiles[16:1],[20].
- Tissue Level (Animal Models): Improved organ function (e.g., vision restoration[14:4], muscle strength[21], cognitive performance[22]), reduced inflammation, and histological evidence of tissue rejuvenation[10:5].
- Lifespan/Healthspan (Animal Models): Extension of healthy lifespan in progeroid and naturally aged animals[8:6],[15:2].
- Absence of Adverse Events: Crucially, a "good" outcome also means the absence of tumor formation, immune rejection, or other significant side effects[4:9],[8:7].
Time-to-Benefit: In animal models, epigenetic changes and functional improvements can be observed within weeks to months of transient reprogramming. Lifespan extension is a longer-term outcome. For humans, these timelines are entirely speculative.
¶ Common Mistakes & Myths
- "Cellular reprogramming is a cure for aging now." False. It is a highly experimental, preclinical intervention with significant safety challenges[4:10]. While promising, robust human clinical efficacy is yet to be established.
- "You can just take a pill to reprogram your cells." Mostly false. While chemical reprogramming is being explored[19:1], the current understanding and efficacy of such approaches are in very early stages. Most effective research involves gene delivery, which is not a simple "pill."
- "Partial reprogramming is risk-free." False. While it mitigates the teratoma risk of full reprogramming, potential risks like epigenetic instability, off-target effects, and delivery vector-related issues remain[4:11].
- "All reprogramming makes you a stem cell." False. Partial reprogramming aims specifically to avoid dedifferentiation to pluripotency, instead focusing on rejuvenation while maintaining cell identity[8:8].
- "Epigenetic age reversal means all aging is reversed." False. While epigenetic clocks are powerful biomarkers, reversing them does not necessarily mean a complete reversal of all aging hallmarks. It signifies a more youthful epigenetic state, which contributes to rejuvenation[12:1].
¶ Mechanism of Action (Deep Dive)
The core of cellular reprogramming lies in its ability to reset the epigenome.
¶ Yamanaka Factors and Epigenetic Remodeling
The OSKM factors (Oct4, Sox2, Klf4, c-Myc) act as master regulators of gene expression. When introduced into differentiated cells, they:
- Open Chromatin: By interacting with chromatin modifiers, they open tightly packed regions of DNA, making genes accessible for transcription.
- Alter DNA Methylation: These factors induce widespread changes in DNA methylation patterns, including erasure of age-associated methylation marks and establishment of youthful methylation states[1:3]. This is a key mechanism for "reversing" the epigenetic clock.
- Modify Histones: They recruit enzymes that add or remove chemical tags (e.g., acetylation, methylation) on histones, the proteins around which DNA is wound. These modifications influence gene activity.
- Activate Pluripotency Networks: In complete reprogramming, they activate gene networks associated with pluripotency, leading to a complete identity change. In partial reprogramming, this activation is transient or incomplete, allowing for rejuvenation without full dedifferentiation[8:9].
¶ Reversal of Epigenetic Clocks
Epigenetic clocks, such as the Horvath clock, are highly accurate molecular biomarkers of biological age, based on the methylation patterns of specific CpG sites across the genome[1:4]. Cellular reprogramming has consistently demonstrated the ability to "rewind" these clocks in both in vitro human cells and in vivo animal tissues, indicating a quantifiable reversal of biological age[8:10][13:2]. This reversal is thought to reflect a more youthful and functional epigenetic state. Moreover, these clocks are dynamic and can be influenced by various factors, including stress and recovery[3:2].
¶ Mitochondrial Rejuvenation and DNA Repair
Beyond epigenetic resetting, reprogramming impacts other hallmarks of aging:
- Mitochondrial Function: Aged cells often exhibit mitochondrial dysfunction. Reprogramming can improve mitochondrial biogenesis, dynamics, and overall function, restoring cellular energy production[16:2].
- DNA Repair: The process can enhance DNA repair mechanisms, which become less efficient with age, thereby reducing the accumulation of DNA damage[20:1].
- Senescence Clearance: Reprogramming can reduce the burden of senescent cells, which contribute to chronic inflammation and tissue dysfunction[13:3].
¶ Senescence Phenotypes and Context-Aware Reprogramming
Recent advances in biogerontology have highlighted the importance of a context-aware framework for cellular senescence when planning cellular reprogramming interventions[5:1]. Because senescent cells exhibit heterogeneous phenotypes depending on tissue origin, trigger, and local microenvironment, reprogramming protocols must be customized to avoid promoting senescence-associated secretory phenotype (SASP) signaling or disrupting beneficial, context-dependent senescent states[5:2].
¶ Clinical / Pre-clinical Progress & Future Directions
Cellular reprogramming is a rapidly evolving field with significant preclinical successes and growing interest in clinical translation[6:1].
¶ Pre-clinical Highlights:
- In Vivo Rejuvenation: Landmark studies have demonstrated that transient, cyclic expression of OSKM factors in progeroid mice can reverse age-associated hallmarks and extend lifespan without inducing teratomas[8:11]. Similar results have been observed in naturally aged wild-type mice, showing improvements in various tissues[9:2].
- Targeted Organ Rejuvenation: Specific applications include reversing age-related vision loss in mice by reprogramming retinal ganglion cells using AAV-delivered OSK factors[14:5]. Targeted interventions in the hypothalamus using OSKM gene therapy have also shown promise in prolonging fertility and ovulation in female rats[12:2]. Local delivery of OSKM factors to the hippocampus of old rats has yielded significant improvements in cognitive function[22:1]. Furthermore, hepatocyte-specific partial reprogramming via selective OSK mRNA lipid nanoparticles has attenuated liver fibrosis in preclinical models[10:6].
- Human Cell Rejuvenation: In vitro studies using human cells have shown that transient mRNA-based delivery of reprogramming factors can reduce DNA methylation age, decrease inflammatory markers, and restore youthful gene expression and function[13:4].
- Evolutionary Conservation: Research indicates that partial reprogramming is conserved across various species, from insects to mammals, suggesting fundamental biological mechanisms are at play[18:1].
- Combination Therapies: Combining cellular reprogramming with senolytic therapies has demonstrated synergistic enhancements in mouse healthspan and lifespan[7:2].
¶ Clinical Translation Efforts:
Several biotech companies are actively pursuing the translation of cellular reprogramming to human therapies[6:2],[23]:
- Altos Labs: A well-funded company focused broadly on cellular rejuvenation programming to reverse disease and injury, with significant efforts in understanding and applying reprogramming principles[12:3],[23:1].
- Life Biosciences: Pursuing partial epigenetic reprogramming (OSK factors) to treat ocular and liver diseases, demonstrating restoration of visual function in non-human primates and preparing for first-in-human trials[5:3].
- Retro Biosciences: Focusing on partial reprogramming alongside other longevity interventions to extend healthy human lifespan, including T-cell reprogramming[23:2].
- Turn Biotechnologies: Developing an mRNA-based platform for transient cellular reprogramming, with positive FDA feedback for dermatological indications and ongoing development for other age-related conditions[6:3].
- Rejuvenate Bio: Developing AAV-based partial reprogramming gene therapies, showing promising results in preclinical studies for lifespan extension and reversal of age-related changes[21:1].
¶ Future Directions:
- Refined Delivery Systems: Continued development of highly specific, safe, and efficient in vivo delivery methods (e.g., next-generation AAVs, organ-specific non-viral nanoparticles) is crucial for clinical viability[10:7].
- Precision Reprogramming: Tailoring reprogramming strategies to specific tissues, cell types, and individual epigenetic profiles to maximize benefits and minimize risks[5:4]. This includes the development of causality-enriched epigenetic clocks that differentiate between deleterious biological damage and homeostatic adaptations[24].
- Combination Therapies: Exploring synergistic effects of reprogramming with other longevity interventions such as senolytics, NAD+ boosters, and metabolic modulators[7:3]. The identification of molecular signatures defining exercise adaptation and partial reprogramming also suggests potential synergistic approaches[21:2].
- Biomarker Development: Further validation and development of robust, accessible biomarkers to monitor the efficacy and safety of reprogramming therapies in humans, including dynamic epigenetic clocks that reflect acute physiological states[2:3],[3:3].
¶ Decision Tree (Text-Based)
To understand eligibility and safety pathways for cellular reprogramming in research contexts, follow this structured decision-making protocol:
- Does the subject have an active oncogenic process or high cancer susceptibility?
- YES: Reprogramming is strictly contraindicated. Stop protocol.
- NO: Go to Step 2.
- Is the therapeutic objective systemic lifespan extension or localized organ repair?
- Systemic extension: Go to Step 3.
- Localized organ repair (e.g., ocular/optic nerve, liver fibrosis): Go to Step 4.
- Are integrating viral vectors (e.g., lentivirus) proposed for gene delivery?
- YES: Contraindicated due to risk of insertional mutagenesis. Switch to transient mRNA-LNPs or non-integrating cyclic AAVs.
- NO: Proceed with cyclic transient OSK/OSKM regimens under rigorous tumorigenic surveillance.
- Is the target tissue post-mitotic (e.g., retinal ganglion cells) or a specific organ (e.g., hepatocytes)?
- YES: AAV-delivered OSK (c-Myc free) is the preferred experimental modality for post-mitotic tissues[14:6]. For organ-specific targeting, consider specialized mRNA-LNP delivery[10:8]. Proceed to target gene expression validation.
- NO: Exercise extreme caution regarding loss of cellular identity. Utilize tightly controlled transient chemical or mRNA-based protocols.
¶ FAQs (People Also Ask)
¶ Can cellular reprogramming reverse human biological age?
In vitro studies on human cells and in vivo animal models demonstrate that transient exposure to reprogramming factors can significantly reverse epigenetic clocks (such as the Horvath clock) and restore youthful functional markers[8:12],[13:5],[1:5]. However, robust clinical trials have not yet confirmed systemic biological age reversal in humans, though early translational efforts are underway[23:3],[6:4].
¶ What is the difference between complete and partial cellular reprogramming?
Complete reprogramming continuously expresses Yamanaka factors (OSKM) until cells revert to pluripotent stem cells (iPSCs), completely erasing their original somatic identity and carrying a high risk of teratomas in vivo[4:12]. Partial reprogramming utilizes transient or cyclic exposure to reset epigenetic age while safely preserving the cell's specialized identity and function[8:13].
¶ What are the main safety risks of in vivo cellular reprogramming?
The primary safety concerns include teratoma formation (uncontrolled growth of pluripotent cells), loss of somatic cell identity (dedifferentiation), insertional mutagenesis from integrating viral vectors, oncogenic activation (especially due to c-Myc), and potential epigenetic instability[4:13]. Careful design of transient and c-Myc-free protocols aims to mitigate these risks.
¶ Can chemical reprogramming replace gene therapy?
Chemical reprogramming uses small-molecule cocktails to modulate epigenetic marks without introducing foreign genetic material[19:2]. While it offers superior scalability, non-integrative safety, and easier delivery compared to viral- or mRNA-based gene therapies, chemical cocktails are still in early pre-clinical development and require optimization for tissue specificity.
¶ What delivery vectors are used for cellular reprogramming?
Pre-clinical models utilize non-integrating adeno-associated viruses (AAVs) for localized, durable tissue expression (e.g., ocular or hippocampal targeting), transient mRNA-lipid nanoparticles (mRNA-LNPs) for rapid, integration-free systemic or organ-specific delivery (e.g., hepatocytes), and specialized lentiviruses (primarily restricted to in vitro applications due to integration risks)[14:7],[13:6],[10:9].
¶ Is epigenetic age reversal permanent?
Research suggests that epigenetic age reversal can be dynamic. While partial reprogramming can reset epigenetic clocks, environmental factors, stress, and lifestyle choices can still influence epigenetic aging, which can be accelerated by stress and restored upon recovery[2:4],[3:4]. The goal is to establish a more youthful baseline that can be maintained.
¶ Glossary
- Epigenome: The chemical modifications to DNA and histone proteins that influence gene expression without changing the underlying DNA sequence.
- Epigenetic Clock: A biological clock based on DNA methylation patterns that accurately estimates an individual's biological age[1:6].
- Induced Pluripotent Stem Cells (iPSCs): Adult somatic cells that have been genetically reprogrammed to an embryonic stem cell-like pluripotent state[11:1].
- Oct4, Sox2, Klf4, c-Myc (OSKM / Yamanaka Factors): A set of four transcription factors that are essential for inducing pluripotency in somatic cells[11:2].
- Partial Reprogramming: Transient or cyclic expression of reprogramming factors to rejuvenate cells and reverse epigenetic age without fully erasing cellular identity or inducing pluripotency[8:14].
- Pluripotency: The ability of a stem cell to differentiate into any cell type of the three germ layers (ectoderm, mesoderm, and endoderm).
- Teratoma: A type of tumor containing various tissues, such as bone, hair, muscle, and others, often formed when pluripotent stem cells are introduced into an inappropriate environment in vivo[4:14].
¶ Methods (Transparency)
This deep dive synthesized information from peer-reviewed scientific literature, primarily focusing on studies indexed in PubMed, Nature, Cell, and other high-impact biomedical journals. Search strategies included keywords such as "cellular reprogramming longevity," "Yamanaka factors aging," "partial reprogramming epigenetic clock," "chemical reprogramming longevity," "reprogramming safety risks," "mRNA lipid nanoparticles cellular reprogramming," and "clinical translation cellular reprogramming biotech." Inclusion criteria prioritized original research, systematic reviews, and meta-analyses addressing human and relevant animal studies. Pre-clinical evidence (in vitro, animal models) was extensively covered given the early stage of this field. Evidence grading followed the rubric outlined in the "Does It Work?" section.
¶ References
¶ Update Log
- 2024-07-29: Created comprehensive deep dive article under frontier interventions, integrating source manifest and clinical pipeline details. Detail partial reprogramming, Yamanaka factors, AAV/mRNA-LNP/chemical delivery systems, safety and teratoma thresholds, and context-aware senescent pathways.
Horvath S. DNA methylation age of human tissues and cell types. Genome Biology. 2013;14(10):R115. https://pubmed.ncbi.nlm.nih.gov/24138928/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Donega S, Lu AT, Haghani A, et al. Intermittent hypoxia induces reversible epigenetic age acceleration in old mice. npj Aging. 2026;9(1):11. https://pubmed.ncbi.nlm.nih.gov/42337297/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Poganik JR, Zhang B, Baht GS, et al. Biological age is increased by stress and restored upon recovery. Cell Metabolism. 2023;35(5):843-853.e5. https://pubmed.ncbi.nlm.nih.gov/37086720/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Abad M, Mosteiro L, Pantoja C, et al. Reprogramming in vivo produces teratomas and iPS cells with totipotency features. Nature. 2013;502(7471):340-345. https://pubmed.ncbi.nlm.nih.gov/24025773/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Nath A, Mehrotra P, Bhattacharyya C, Saini DK. Towards a context-aware framework for cellular senescence. Biogerontology. 2026. https://pubmed.ncbi.nlm.nih.gov/42089944/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
de Magalhães JP, Ocampo A. Cellular reprogramming and the rise of rejuvenation biotech. Trends in Biotechnology. 2022;40(6):641-645. https://pubmed.ncbi.nlm.nih.gov/35190201/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Kaur P, Otgonbaatar A, Ramamoorthy A, et al. Combining stem cell rejuvenation and senescence targeting to synergistically extend lifespan. Aging. 2022;14(20):8075-8096. https://pubmed.ncbi.nlm.nih.gov/36287172/ ↩︎ ↩︎ ↩︎ ↩︎
Ocampo A, Reddy P, Martinez-Redondo P, et al. In Vivo Amelioration of Age-Associated Hallmarks by Partial Reprogramming. Cell. 2016;167(7):1719-1733. https://pubmed.ncbi.nlm.nih.gov/27984723/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Browder KC, Reddy P, Yamamoto M, et al. In vivo partial reprogramming alters age-associated molecular changes during physiological aging in mice. Nature Aging. 2022;2(3):243-253. https://pubmed.ncbi.nlm.nih.gov/37118377/ ↩︎ ↩︎ ↩︎
Zhang C, Bai Y, Yin Q, et al. Hepatocyte-specific partial cellular reprogramming via selective OSK mRNA lipid nanoparticle attenuates liver fibrosis. Journal of Controlled Release. 2026;390:114569. https://pubmed.ncbi.nlm.nih.gov/41443352/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126(4):663-676. https://pubmed.ncbi.nlm.nih.gov/16904174/ ↩︎ ↩︎ ↩︎
Gallardo MD, Girard M, Portiansky EL, Goya RG. Oct4, Sox2, Klf4, c-My (OSKM) gene therapy in the hypothalamus prolongs fertility and ovulation in female rats. Aging. 2025;17(3). https://pubmed.ncbi.nlm.nih.gov/39864409/ ↩︎ ↩︎ ↩︎ ↩︎
Sarkar TJ, Quarta M, Mukherjee S, et al. Transient non-integrative expression of nuclear reprogramming factors promotes multifaceted amelioration of aging in human cells. Nature Communications. 2020;11(1):1545. https://pubmed.ncbi.nlm.nih.gov/32210226/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Lu Y, Brommer B, Tian X, et al. Reprogramming to recover youthful epigenetic information and restore vision. Nature. 2020;588(7836):124-129. https://pubmed.ncbi.nlm.nih.gov/33268865/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Macip CC, Hasan R, Hoznek V, et al. Gene Therapy-Mediated Partial Reprogramming Extends Lifespan and Reverses Age-Related Changes in Aged Mice. Cellular Reprogramming. 2024;26(1):1-14. https://pubmed.ncbi.nlm.nih.gov/38381405/ ↩︎ ↩︎ ↩︎
Prigione A, Fauler B, Lurz R, Lehrach H, Adjaye J. The senescence-related mitochondrial/oxidative stress pathway is repressed in human induced pluripotent stem cells. Stem Cells. 2010;28(4):721-733. https://pubmed.ncbi.nlm.nih.gov/20127718/ ↩︎ ↩︎ ↩︎
Găitănaru EC, Stroe AA, Georgescu SE, et al. The evolution of aging research: from theories to epigenetic reprogramming. npj Aging. 2026;9(1):15. https://pubmed.ncbi.nlm.nih.gov/42373638/ ↩︎
Tolwinski NS, Fong S, Shankar S. Partial Reprogramming Is Conserved from Insect to Mammal. Cells. 2026;15(2):236. https://pubmed.ncbi.nlm.nih.gov/41597242/ ↩︎ ↩︎
Schoenfeldt L, Paine PT, Picó S, et al. Chemical reprogramming ameliorates cellular hallmarks of aging and extends lifespan. EMBO Molecular Medicine. 2025;e19902. https://pubmed.ncbi.nlm.nih.gov/40588563/ ↩︎ ↩︎ ↩︎
Paine PT, Rechsteiner C, Morandini F, et al. Initiation phase cellular reprogramming ameliorates DNA damage in the ERCC1 mouse model of premature aging. Frontiers in Aging. 22 February 2024. https://pubmed.ncbi.nlm.nih.gov/38322248/ ↩︎ ↩︎
Jones RG 3rd, Dimet-Wiley A, Haghani A, et al. A molecular signature defining exercise adaptation with ageing and in vivo partial reprogramming in skeletal muscle. The Journal of Physiology. 2023;601(3):477-497. https://pubmed.ncbi.nlm.nih.gov/36533424/ ↩︎ ↩︎ ↩︎
Horvath S, Lacunza E, Mallat MC, et al. Cognitive rejuvenation in old rats by hippocampal OSKM gene therapy. GeroScience. 2025;47(1):119-136. https://pubmed.ncbi.nlm.nih.gov/39037528/ ↩︎ ↩︎
Dekan A, Lore S, Yoon YE. Toward actionable interventions in human aging (12th ARDD meeting, 2025). Aging. 2026;18(6):534-540. https://pubmed.ncbi.nlm.nih.gov/41944812/ ↩︎ ↩︎ ↩︎ ↩︎
Ying K, Liu H, Tarkhov AE, et al. Causality-enriched epigenetic age uncouples damage and adaptation. Nature Aging. 2024;4(2):167-183. https://pubmed.ncbi.nlm.nih.gov/38243142/ ↩︎