¶ Gut Barrier
The gut barrier represents the vast, highly specialized physical and immunological interface that separates the host's internal environment from the external milieu of the gastrointestinal lumen. Spanning approximately 30 to 40 square meters, this multi-layered defensive system is task-balanced with opposing roles: it must remain selectively permeable to facilitate nutrient, water, and electrolyte absorption, while simultaneously presenting an impenetrable block against pathogenic microbes, dietary antigens, and environmental toxins.
¶ At a Glance
¶ Key Points
- Multi-layered defense: The gut barrier is not a single wall but an integrated system consisting of physical, cellular, and immunological layers.
- Microbial symbiosis: The outermost layer of the barrier consists of the commensal microbiota, which actively repels pathogens and processes metabolic inputs.
- Physical monolayer: A single layer of specialized epithelial cells (enterocytes) forms the core physical boundary, sealed by tight junctions.
- Immune surveillance: Over 70% of the host's immune cells reside immediately beneath the epithelial lining, providing rapid response capacity.
- Age-associated decline: Natural aging processes lead to mucus thinning, dysbiosis, and compromised structural integrity, contributing to systemic low-grade inflammation (inflammaging).
¶ Primary Functions
- Selective absorption: Transports amino acids, lipids, carbohydrates, vitamins, and minerals.
- Pathogen exclusion: Blocks translocation of lipopolysaccharides (LPS), bacteria, fungi, and viruses.
- Immune tolerance: Prevents hyper-inflammatory responses to benign dietary proteins and symbiotic commensals.
¶ Anatomical Structure & Physiology
The intestinal barrier is a sophisticated, progressive assembly of biological defenses that operate in concentric layers from the lumen to the bloodstream.
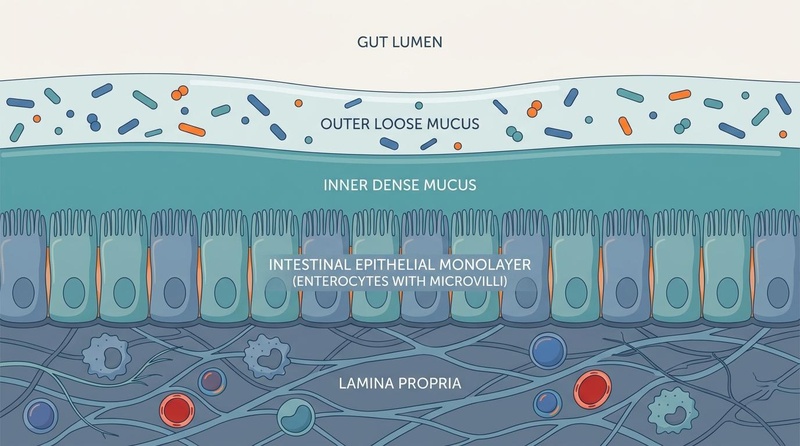
Figure 1: The Multi-Layered Architecture of the Gut Barrier, illustrating the symbiotic relationship between mucosal defenses, the epithelial monolayer, and mucosal immunity.
¶ 1. The Luminal & Microbiota Layer
The outermost functional layer of the gut barrier resides within the lumen.
- Commensal Microbiota: Trillions of symbiotic microorganisms (primarily Bacteroidetes, Firmicutes, and Actinobacteria) outcompete opportunistic pathogens for nutrients and mucosal attachment sites. This phenomenon, known as colonization resistance, prevents pathogen colonization.
- Metabolic Processing: Commensal bacteria ferment dietary fibers to produce short-chain fatty acids (SCFAs) like acetate, propionate, and butyrate. Butyrate serves as the primary energy source for colonocytes and is essential for maintaining barrier transcription factors [1].
- Antimicrobial Secretion: Commensal species synthesize bacteriocins and other antimicrobial peptides that selectively target invading pathogens.
¶ 2. The Dual Mucus Layer
The mucus layer serves as the primary physical barrier preventing direct contact between luminal contents and the delicate epithelial cell layer.
- Outer Loose Mucus Layer: Populated by commensal microbes, this layer is less dense and provides a habitat for symbiotic species that utilize mucin glycans as energy sources.
- Inner Dense Mucus Layer: Virtually sterile and highly compacted, this layer is firmly attached to the epithelial cells. It consists of highly glycosylated polymeric mucin-2 (MUC2) networks secreted by specialized goblet cells. The pore size of this network is smaller than most bacteria, physically excluding them from the cellular surface.
¶ 3. The Intestinal Epithelial Monolayer
A single layer of columnar epithelial cells forms the primary physical boundary of the host. These cells are continually renewed every 3 to 5 days from pluripotent stem cells located in the crypts of Lieberkühn.
- Enterocytes: Representing over 80% of the mucosal cells, these polarized cells feature apical microvilli (the brush border) designed to optimize nutrient absorption.
- Goblet Cells: Specialized secretory cells responsible for the synthesis, packaging, and exocytosis of mucin-2 glycans, maintaining the protective mucus layers.
- Paneth Cells: Located at the base of the crypts, these cells secrete essential antimicrobial proteins (AMPs) including alpha-defensins, lysozyme, and secretory phospholipase A2 (sPLA2) to keep the crypt environment sterile.
- Enteroendocrine Cells: Sensory cells that release regulatory peptides and hormones (e.g., GLP-1, GLP-2, PYY, serotonin) in response to luminal nutrient sensing.
¶ 4. The Lamina Propria & Mucosal Immune Layer
Directly beneath the basement membrane lies the lamina propria, a loose connective tissue layer containing the gut-associated lymphoid tissue (GALT).
- Secretory IgA (sIgA): Plasma cells in the lamina propria produce dimeric IgA, which is actively transported across the enterocytes into the mucus layer. Here, sIgA neutralizes toxins and pathogens via immune exclusion without triggering inflammatory cascades.
- Immune Cell Network: Dendritic cells, macrophages, T-lymphocytes (including regulatory T-cells or Tregs), and B-cells monitor translocated antigens. Dendritic cells can project dendrites between enterocytes directly into the lumen to sample antigens without disrupting tight junction integrity.
¶ Molecular Mechanisms of Regulation
The structural integrity of the epithelial monolayer is maintained by the apical junctional complex (AJC), which regulates the paracellular pathway (the space between adjacent cells).
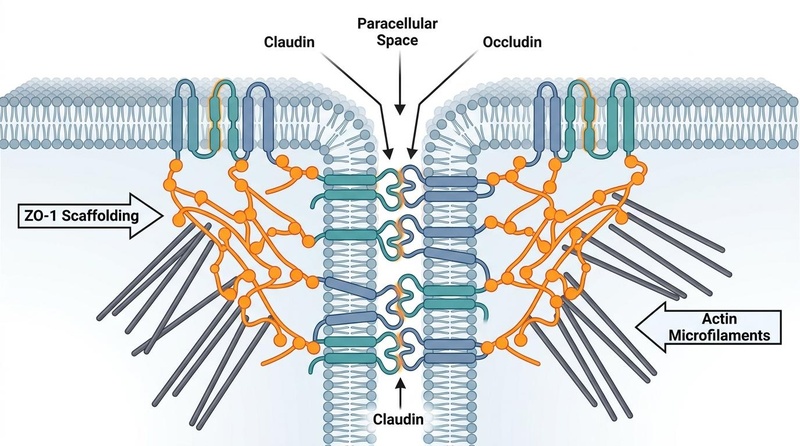
Figure 2: Molecular Architecture of the Tight Junction (Zonula Occludens) Complex. Transmembrane proteins claudins and occludin seal the paracellular space, anchored internally by scaffolding proteins (ZO-1, ZO-2, ZO-3) connected directly to the perijunctional actin-myosin cytoskeleton.
The junctional complex consists of three distinct zones, ordered from apical to basal:
¶ 1. Tight Junctions (Zonula Occludens)
These are the primary regulators of paracellular permeability, forming a continuous band around the apical pole of each epithelial cell.
- Transmembrane Barrier Proteins:
- Claudins: A family of proteins that determine the charge- and size-selectivity of the paracellular pore. "Barrier-forming" claudins (e.g., Claudin-1, -3, -4, -5, -8) seal the junction, whereas "pore-forming" claudins (e.g., Claudin-2, -15) create selective channels for water and small ions.
- Occludin: Regulates macromolecular paracellular flux and stabilizes the tight junction assembly.
- Junction Adhesion Molecules (JAMs): Immunoglobulin-superfamily proteins involved in cell-to-cell adhesion and regulating leukocyte extravasation.
- Intracellular Scaffolding Proteins:
- Zonula Occludens (ZO-1, ZO-2, ZO-3): Multi-domain adapter proteins that link the cytoplasmic tails of claudins and occludin directly to the perijunctional actomyosin ring (PAMR). ZO-1 is critical for coordinating the structural assembly of the tight junction.
¶ 2. Adherens Junctions (Zonula Adherens)
Located below the tight junctions, adherens junctions are critical for cell-to-cell adhesion and structural stability. They are composed of transmembrane E-cadherin proteins that bind homophilically to E-cadherin on adjacent cells, anchored intracellularly by alpha-, beta-, and gamma-catenins to actin filaments. Adherens junctions provide the mechanical tension required for tight junctions to seal properly.
¶ 3. Desmosomes (Macula Adherens)
Located basally, desmosomes resist mechanical shearing forces, anchoring the intermediate filaments (keratin) of the cytoskeleton to the plasma membrane via desmogleins and desmocollins.
¶ Physiological Interactions (The Systemic Axes)
The gut barrier does not operate in isolation; it functions as a critical node in several systemic bidirectional axes.
¶ The Gut-Brain-Microbiome Axis
Bi-directional communication occurs via direct neural pathways (primarily the vagus nerve), endocrine signaling (gut hormones), and microbial metabolites [2].
- Epithelial Integrity and Mood: Compromised barrier function allows bacterial cell-wall components like Lipopolysaccharides (LPS) to enter the portal vein, triggering low-grade systemic inflammation. This neuroinflammatory signaling alters microglia activation and central neurotransmitter synthesis, contributing to cognitive decline and mood disorders [3].
- Autonomic Regulation: Efferent vagal signals can suppress mucosal inflammation and preserve tight junction expression via the cholinergic anti-inflammatory pathway.
¶ The Gut-Liver Axis
The liver receives approximately 75% of its blood supply directly from the gastrointestinal tract via the portal vein, making it the second line of defense against translocated gut antigens.
- First-Pass Filtration: Under physiological conditions, the liver's resident macrophages (Kupffer cells) clear trace amounts of translocated bacterial products from the portal circulation.
- Hepatic Pathology: If the gut barrier fails, a flood of pathogen-associated molecular patterns (PAMPs) reaches the liver. This activates hepatic stellate cells and Kupffer cells via TLR4 pathways, driving the progression of metabolic dysfunction-associated steatotic liver disease (MASLD) and hepatic fibrosis [4].
¶ The Gut-Kidney Axis
Renal function and intestinal barrier integrity are highly interdependent [5].
- Uremic Toxins: In renal impairment, retained uremic toxins (such as indoxyl sulfate and p-cresol sulfate) accumulate in the circulation. These toxins diffuse into the gut lumen, where they are converted by dysbiotic microbes into ammonia, which direct-toxicity damages tight junctions and impairs enterocyte mitochondrial function [5:1].
- Mitochondrial Support: Intestinal SCFAs support renal tubular cell mitochondrial respiration, showing that protecting the gut barrier preserves renal microvascular homeostasis [5:2].
¶ Age & Sex Differences in Barrier Homeostasis
Gut barrier structure and performance undergo systematic changes across the lifespan and differ significantly between sexes.
¶ Lifespan Trajectory & Older Adults
- Mucus Layer Thinning: With advanced age, the expression of MUC2 decreases, and goblet cell secretory capacity declines. This thins the inner dense mucus layer, bringing luminal microbes into direct contact with epithelial cells.
- Immunosenescence of the GALT: Older adults exhibit a decline in secretory IgA (sIgA) diversity and a down-regulation of antimicrobial peptides (AMPs) from Paneth cells.
- Tight Junction Redistribution: Aging is associated with a baseline increase in pore-forming Claudin-2 and a decrease in barrier-forming Claudin-1, leading to a state of chronic, low-grade mucosal leakiness that feeds systemic inflammaging [6].
¶ Sex-Specific Differences
- Estrogen Protection: Estrogens, particularly 17β-estradiol (E2), exert a protective effect on the gut barrier. E2 binds to estrogen receptor beta (ERβ) on enterocytes, directly upregulating the transcription of occludin and junctional adhesion molecules.
- Permeability Fluctuations: Premenopausal women often experience minor cyclic changes in gut permeability corresponding to fluctuations in estradiol levels during the menstrual cycle.
- Postmenopausal Shift: Following menopause, the decline in circulating E2 and the down-regulation of ERβ signaling lead to a rapid increase in baseline intestinal permeability and a shift toward an inflammatory mucosal immune profile [7].
¶ Biomarkers & Clinical Measurement
Assessing intestinal barrier function in vivo requires distinct, validated biomarkers that isolate specific layers of the barrier.
| Biomarker | Specimen | Target | Clinical Interpretation | Key Citations |
|---|---|---|---|---|
| Fecal Zonulin | Stool | Tight Junction Regulation | Elevated levels indicate active disassembly of tight junctions. Primarily triggered by gliadin or dysbiosis. | [8][9] |
| Fecal Calprotectin | Stool | Mucosal Inflammation | A calcium-binding protein from neutrophils. High levels confirm active mucosal inflammation and structural damage. | [10] |
| LPS-Binding Protein (LBP) | Serum | Systemic Endotoxemia | LBP binds to translocated bacterial lipopolysaccharides. Elevated serum LBP indicates chronic, systemic bacterial translocation. | [5:3] |
| Secretory IgA (sIgA) | Stool | Mucosal Immunity | Measures the secretory immune capacity of the GALT. Low levels suggest depleted local defense and increased risk of pathogen binding. | [11] |
| Lactulose/Mannitol Ratio | Urine | Paracellular vs. Transcellular Flux | Dual-sugar absorption test. High recovery ratio in urine indicates increased paracellular passage of the large sugar (lactulose). | [12] |
¶ Biomarker Detail & Limitations
- Zonulin: Zonulin is the human protein analogue of the cholera toxin Zonula occludens toxin (Zot). It is released by enterocytes in response to CXCR3 receptor activation by gliadin or bacterial overgrowth [13]. Once secreted, zonulin binds to epidermal growth factor receptors (EGFR) and protease-activated receptor 2 (PAR2) on adjacent cells, triggering a PLC/PKC-dependent pathway that polymerizes actin and pulls tight junctions apart [8:1][14].
- LPS vs. LBP: Measuring free serum LPS is technically challenging due to its short half-life and environmental contamination risks. Measuring serum LBP provides a highly stable, long-term surrogate marker of systemic endotoxemic exposure.
¶ References
Wang, M., et al. (2026). "Short-chain fatty acids of intestinal origin attenuate protein-bound uremic toxins in patients with chronic kidney disease by protecting the intestinal barrier: a pooled analysis." BMC Gastroenterology, 26(1), 112. https://pubmed.ncbi.nlm.nih.gov/42289655/ ↩︎
Cryan, J. F., et al. (2019). "The microbiota-gut-brain axis." Physiological Reviews, 99(4), 1877-2013. https://pubmed.ncbi.nlm.nih.gov/31460832/ ↩︎
Turek, A., et al. (2026). "Serum LBP and zonulin levels with brain MRS findings and gastrointestinal symptoms in schizophrenia and health." Brain, Behavior, & Immunity - Health, 37, 100762. https://pubmed.ncbi.nlm.nih.gov/41852948/ ↩︎
Chen, Y., et al. (2026). "Gut microbiota in type 2 diabetes mellitus: mechanistic links between dysbiosis, insulin resistance, and chronic low-grade inflammation." Frontiers in Endocrinology, 17, 895. https://pubmed.ncbi.nlm.nih.gov/42375324/ ↩︎
Hu, Q., et al. (2026). "The role of mitochondria in the gut-kidney axis: implications for kidney health." Frontiers in Pharmacology, 17, 1045. https://pubmed.ncbi.nlm.nih.gov/42394968/ ↩︎ ↩︎ ↩︎ ↩︎
López-Otín, C., et al. (2023). "Hallmarks of aging: An expanding universe." Cell, 186(2), 243-278. https://pubmed.ncbi.nlm.nih.gov/36599349/ ↩︎
Tume, R., et al. (2026). "Sex differences in the associations between lifestyle, intestinal permeability and brain health in middle-aged adults." Brain, Behavior, & Immunity - Health, 38, 100780. https://pubmed.ncbi.nlm.nih.gov/42294080/ ↩︎
Fasano, A. (2020). "All disease begins in the (leaky) gut: role of zonulin-mediated gut permeability in the pathogenesis of some chronic inflammatory diseases." F1000Research, 9, F1000 Faculty Rev-98. https://pubmed.ncbi.nlm.nih.gov/32051759/ ↩︎ ↩︎
Tok, A. C., & Sayın, O. (2026). "Serum Zonulin and Chitinase (CHI3L1) as Biomarkers of Intestinal Permeability and Disease Activity in Pediatric Celiac Disease." Children, 13(5), 390. https://pubmed.ncbi.nlm.nih.gov/42353900/ ↩︎
Frandeș, S. I., et al. (2026). "Fecal Zonulin-Related Proteins in Inflammatory Bowel Disease: Associations with Clinical Disease Activity and Inflammatory Markers." Medicina, 62(6), 156-168. https://pubmed.ncbi.nlm.nih.gov/42356172/ ↩︎
Ion, L. M., et al. (2026). "Integrated Immune-Gut Profiling Identifies an Exploratory Pediatric Inflammatory Intestinal Profile Associated with Food-Specific IgG Reactivity." Biomolecules, 16(6), 423. https://pubmed.ncbi.nlm.nih.gov/42352388/ ↩︎
Camilleri, M. (2021). "Human Intestinal Barrier: Effects of Stressors, Diet, Prebiotics, and Probiotics." Clinical and Translational Gastroenterology, 12(1), e00308. https://pubmed.ncbi.nlm.nih.gov/33492118/ ↩︎
Lammers, K. M., et al. (2008). "Gliadin induces an increase in intestinal permeability and zonulin release by binding to the chemokine receptor CXCR3." Gastroenterology, 135(1), 194-204. https://pubmed.ncbi.nlm.nih.gov/18485912/ ↩︎
Sturgeon, C., & Fasano, A. (2016). "Zonulin, a regulator of epithelial and endothelial barrier functions, and its involvement in chronic inflammatory diseases." Tissue Barriers, 4(4), e1251384. https://pubmed.ncbi.nlm.nih.gov/28123927/ ↩︎
See also: Leaky Gut (Intestinal Permeability), Dysbiosis