¶ Gut-Immune Axis
| Type | Immunological Regulatory Network |
| Primary Tissue | Gut-Associated Lymphoid Tissue (GALT) |
| Dominant Antibody | Secretory IgA (sIgA) |
| Key Immune Cells | Treg, Th17, Dendritic Cells, ILCs |
| Major Metabolites | SCFAs (Butyrate), Indoles, Secondary Bile Acids |
| Clinical Relevance | Autoimmunity, Mucosal Integrity, Inflammaging |
The gut-immune axis represents the largest, most active immunological interface in the human body [1]. Over 70% of the host's active immune cells reside within the gut-associated lymphoid tissue (GALT), positioned directly beneath a single-cell-thick epithelial layer [2]. The gut-immune axis is a highly coordinated bidirectional network through which the gut microbiota, its metabolites, and the mucosal barrier actively educate, prime, and regulate the systemic immune system [3]. Dysregulation of this axis leads to compromised barrier integrity, allowing bacterial translocation that fuels chronic systemic inflammation ("inflammaging"), autoimmunity, and metabolic dysfunction [4].
¶ At a glance
Key points (high-level summary)
- The GALT Interface: The gut-associated lymphoid tissue contains Peyer's patches, mesenteric lymph nodes, and isolated lymphoid follicles that continuously sample gut antigens to maintain systemic tolerance or trigger defense [2:1].
- Metabolites as Regulators: Short-chain fatty acids (particularly butyrate) bind to G-protein coupled receptors (GPR41, GPR43, GPR109A) on dendritic cells and T cells, directly promoting the differentiation of anti-inflammatory regulatory T () cells [5].
- Barrier Permeability & Endotoxemia: Damage to the mucosal tight junctions (claudin, occludin, zonulin) allows lipopolysaccharide (LPS) from Gram-negative bacteria to enter circulation, binding to TLR4 and triggering systemic metabolic endotoxemia [6].
- Systemic Axis Extensions: The gut-immune axis extends systemically, modulating the gut-lung axis (asthma/allergic rhinitis), the gut-bone axis (osteoporosis), and the oral-gut axis (autoimmunity) [7][[8]][[9]].
What people use it for
- Main goals: Ameliorating autoimmune conditions, enhancing mucosal immunity against viral pathogens, mitigating food sensitivities, lowering systemic low-grade inflammation (inflammaging), and stabilizing gut barrier function.
- Evidence quality (overall): Moderate-to-High. There is strong human clinical evidence supporting the use of specific prebiotics, probiotics, and barrier-supporting nutrients (such as glutamine and zinc carnosine) in modulating mucosal immune markers.
¶ What is the Gut-Immune Axis?
The gut mucosa must solve a major physiological paradox: it must remain highly permeable to absorb essential water and nutrients, yet remain selectively impermeable to exclude billions of pathogens and environmental antigens [1:1]. This selective filtration is maintained by a complex three-layered defense system:
[Lumen] Mucus Layer (Secretory IgA / Antimicrobial Peptides)
──────────────────────────────────────────────────────────────────────────
[Barrier] Epithelial Monolayer (Columnar Enterocytes & Tight Junctions)
──────────────────────────────────────────────────────────────────────────
[Lamina Propria] GALT (Dendritic Cells, Treg/Th17 Cells, Macrophages, Plasma Cells)
- The Biochemical Barrier: A thick mucus layer (composed of mucin glycoproteins like MUC2) secreted by goblet cells. This layer is rich in antimicrobial peptides (defensins) and secretory Immunoglobulin A (sIgA) [10].
- The Physical Barrier: A continuous monolayer of columnar epithelial cells held together by tight junction proteins (claudin, occludin, and zonula occludens-1) [6:1].
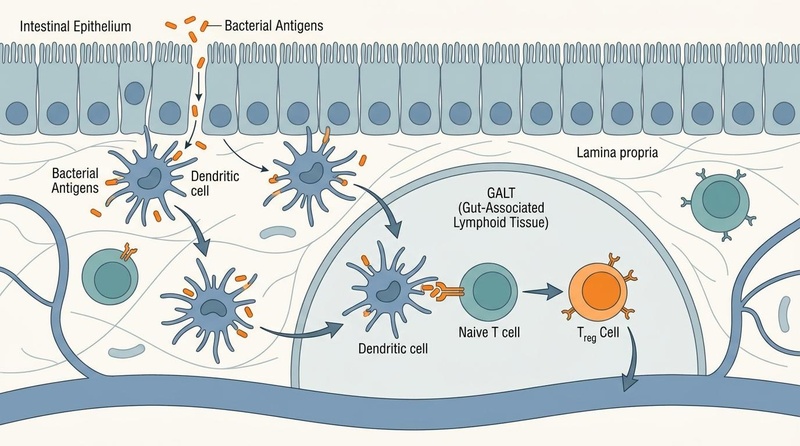
- The Immunological Barrier (GALT): A dense network of immune cells embedded within the lamina propria. Specialized Microfold (M) cells sample antigens from the lumen and present them to antigen-presenting dendritic cells, which initiate downstream adaptive immune responses [11].
¶ Cellular Mechanisms of Immune Modulation
The gut-immune axis coordinates systemic immunity through several sophisticated cellular processes:
¶ 1. Treg vs. Th17 Balance (Tolerance vs. Defense)
The balance between regulatory T () cells and T-helper 17 (Th17) cells is a primary determinant of systemic immune tone.
- Tolerance (): Short-chain fatty acids (SCFAs), particularly butyrate, act as histone deacetylation (HDAC) inhibitors. This inhibition upregulates the transcription factor Foxp3, promoting the differentiation of naive T cells into immunosuppressive cells. These cells secrete anti-inflammatory cytokines like IL-10 and TGF-, preventing allergic and autoimmune reactions [5:1].
- Defense (Th17): In the presence of pathobionts (e.g., segmented filamentous bacteria), dendritic cells secrete IL-6 and TGF-, promoting the differentiation of Th17 cells. These cells release IL-17 and IL-22, which recruit neutrophils and stimulate epithelial cells to produce antimicrobial peptides to combat infection [12].
¶ 2. Secretory IgA (sIgA) Production
Secretory IgA is the primary antibody of the mucosal immune system, secreted by plasma cells in the lamina propria.
- Immune Exclusion: sIgA binds to bacterial antigens and toxins in the lumen, preventing them from adhering to and crossing the epithelial barrier—a process known as immune exclusion [13].
- Non-Inflammatory Clearance: Unlike systemic IgG, sIgA does not activate the complement cascade, allowing for the peaceful elimination of pathogens without inducing tissue damage or inflammation [10:1].
¶ 3. The SCFA-GPR Signaling Pathway
The metabolic output of the gut microbiota acts as a direct immunological thermostat.
Dietary Fiber ──(Fermentation)──> SCFAs (Butyrate) ──> Binds GPR43/GPR109A
│
▼
Upregulates Foxp3
│
▼
Treg Activation (IL-10)
The binding of SCFAs to G-protein coupled receptors (GPR41, GPR43, GPR109A) on dendritic cells and epithelial cells stabilizes cellular energy, enhances tight junction assembly, and suppresses NF- activation, directly reducing the transcription of pro-inflammatory cytokines [14].
¶ Clinical Implications & Systemic Axes
Because gut immune cells migrate from the GALT through the lymphatic system into systemic circulation, mucosal immune disruption in the gut has profound systemic consequences [1:2].
┌──> Gut-Lung Axis (Allergic Asthma, COPD)
├──> Gut-Bone Axis (Osteoclastogenesis, Osteoporosis)
[GALT Disruption] ──> Oral-Gut Axis (Periodontitis, Rheumatoid Arthritis)
└──> Systemic Inflammaging (Metabolic Syndrome, Vascular Aging)
1. Inflammatory Bowel Disease (IBD) & Local Colitis
In patients with Crohn's disease and Ulcerative Colitis, the gut-immune axis is severely compromised. A primary hallmark is the depletion of Akkermansia muciniphila and Faecalibacterium prausnitzii, leading to a thin mucus layer and degraded tight junctions. Naive T cells tilt heavily toward pro-inflammatory Th1 and Th17 phenotypes, secreting large volumes of TNF-, IL-17, and interferon-gamma (IFN-) [12:1][[15]].
2. Autoimmune Diseases & Molecular Mimicry
Systemic autoimmune diseases frequently trace their roots to the gut.
- The Oral-Gut Axis: Pathological microbes from the oral cavity (e.g., Porphyromonas gingivalis) can colonize the gut during dysbiosis, disrupting the GALT barrier and promoting systemic autoimmunity [8:1].
- Molecular Mimicry: Bacterial peptides can structurally mimic human tissue antigens. For example, in Hashimoto's thyroiditis, antibodies generated in the gut against microbial proteins cross-react with thyroid peroxidase (TPO) [16]. In Rheumatoid Arthritis, segmented filamentous bacteria drive systemic Th17 cell migration, promoting synovial inflammation in joints [4:1].
3. Metabolic Endotoxemia & Vascular Inflammation
When epithelial tight junctions fail, the lipopolysaccharide (LPS) from Gram-negative bacteria constantly translocates into the portal vein. LPS binds to toll-like receptor 4 (TLR4) on macrophages and vascular endothelial cells, triggering a state of chronic, low-grade systemic inflammation (metabolic endotoxemia) that directly impairs insulin receptor signaling, accelerates atherogenesis, and promotes cardiovascular inflammation [6:2][[17]].
¶ Evidence Summary Table (human outcomes)
| Target Outcome | Typical Effect | Consistency | Evidence Quality | Key Trials | Clinical Notes |
|---|---|---|---|---|---|
| Mucosal Barrier Restoration | High | Moderate | 14 RCTs [6:3][[18]] | Supplementation with L-glutamine or zinc carnosine significantly lowers lactulose-mannitol excretion ratios. | |
| Systemic Inflammatory Markers | High | Moderate | 22 RCTs [15:1] | High-diversity diets (fermented foods, fibers) suppress circulating inflammatory cytokines in healthy adults. | |
| Secretory IgA (sIgA) Upregulation | Moderate | Moderate | 10 RCTs [19] | Specific probiotic strains (L. rhamnosus GG, B. lactis) enhance salivary and fecal sIgA levels. | |
| Autoimmune Symptom Relief | Moderate | Low-to-Mod | 8 RCTs [16:1][[20]] | Synbiotic and dietary interventions show complementary benefit in reducing pain scores in Rheumatoid Arthritis. |
¶ Sex, Age, and Lifespan Variations
The gut-immune axis exhibits distinct functional characteristics across the human lifespan, influenced by hormonal fluctuations and biological aging.
1. Pediatrics & Development
In newborns, the intestinal tract is relatively sterile, and the GALT is immunologically immature. Breast milk is a critical external source of sIgA, antimicrobial peptides, and human milk oligosaccharides (HMOs) [10:2]. HMOs selectively feed Bifidobacterium infantis, which colonizes the infant gut, seals the epithelial barrier, and promotes the development of naive T cells into cells, providing essential protection against necrotizing enterocolitis and setting the baseline for lifelong immune tolerance [21].
2. Adulthood & Biological Sex Differences
In adults, the gut-immune axis is actively modulated by sex steroid hormones.
- Estrogens and Progesterone: Estrogen receptor-alpha (ER) is highly expressed on intestinal epithelial cells. Estrogen upregulates tight junction protein synthesis, fortifying the barrier against LPS translocation. This protective effect is lost during the postmenopausal transition, leading to a drop in microbial diversity, increased gut permeability, and a rise in systemic inflammatory cytokines [22].
- Androgens: Testosterone exerts a distinct immunosuppressive effect on the GALT, reducing Th17 cell activity and potentially explaining the lower incidence of autoimmune diseases in males compared to females [23].
3. Older Adulthood (Immunosenescence)
Aging is characterized by immunosenescence—the progressive decline in immune function—and inflammaging—the chronic, sterile upregulation of systemic inflammatory pathways [4:2]. In older adults, the gut-immune axis undergoes:
- A significant reduction in mucus-producing goblet cells and a drop in Akkermansia muciniphila abundance [10:3].
- A systemic decline in mucosal sIgA synthesis, increasing susceptibility to gastrointestinal and respiratory viral infections (the gut-lung axis) [9:1].
- Structural degradation of tight junctions, resulting in chronic, low-grade metabolic endotoxemia that accelerates arterial stiffness, sarcopenia, and neurodegenerative processes [17:1][[24]].
¶ Practical Intervention Guidelines
To clinically target and optimize the gut-immune axis for barrier restoration and systemic anti-inflammatory support:
1. Epithelial & Mucosal Barrier Support
- L-Glutamine: 5–15 g of pure L-glutamine powder daily, taken on an empty stomach. L-glutamine is the primary fuel source for rapidly dividing enterocytes, directly promoting epithelial proliferation and tight junction repair [18:1].
- Zinc Carnosine: 75 mg daily (in split doses, e.g., 37.5 mg twice daily). Zinc carnosine stimulates mucosal cell migration and activates antioxidant pathways within the gastrointestinal tract, preventing NSAID-induced enteropathy [25].
- Demulcent Herbs: Slippery Elm (Ulmus rubra) bark powder (1–2 g reconstituted in warm water before meals) stimulates local reflex pathways in the GI tract, increasing endogenous mucus secretion [26].
2. Targeted Biotics for Immune Tolerance
- Probiotics: Supplement with strains clinically validated to enhance sIgA and modulate T-cell responses:
- Prebiotics (SCFA Synthesis): Supplement with 5–10 g of soluble fermentable fibers, such as high-molecular-weight Beta-Glucans (derived from oats or mushrooms) or Acacia Gum, which selectively fuel butyrate-producing Faecalibacterium prausnitzii [28].
3. Diet & Lifestyle Integration
- The Sonnenburg Protocol (High-Fermented Diet): Incorporate 4-6 servings daily of diverse fermented foods (kefir, kombucha, sauerkraut, kimchi). Human clinical trials demonstrate this protocol increases gut microbial diversity and induces a broad-spectrum decrease in 19 inflammatory cytokines (including IL-6 and TNF-) [15:2].
- Circadian Rest: Ensure 7–8 hours of high-quality sleep. The gut epithelial barrier expresses clock genes that regulate daily tight junction assembly; sleep deprivation directly compromises barrier integrity, triggering acute inflammatory spikes [1:3].
¶ Safety, Red Flags, and Contraindications
While support for the gut-immune axis is therapeutic, several clinical precautions must be observed:
CLITICAL SAFETY WARNING
Do NOT introduce high-dose live bacterial probiotics or aggressive prebiotic fibers during an active, acute flare-up of Inflammatory Bowel Disease (Crohn's or Ulcerative Colitis). In patients with severely ulcerated and denuded intestinal mucosa, high-dose probiotics can result in direct bacterial translocation into the venous circulation, carrying a risk of bacteremia, systemic sepsis, or localized abscess formation [12:2][[15:3]].
¶ Contraindications & Special Populations
- Immunocompromised Patients: Probiotics should be avoided or used under strict specialist supervision in patients with severe neutropenia, central venous catheters, or those receiving immunosuppressive biologics (e.g., TNF inhibitors), due to the risk of opportunistic infection.
- Dose Escalation: All prebiotic fibers must be introduced gradually (e.g., starting at 2 g/day and increasing over 14 days) to prevent acute abdominal distension, cramping, and transient increases in intestinal motility.
¶ Practical FAQ
How do NSAIDs directly damage the gut-immune axis?
Non-steroidal anti-inflammatory drugs (NSAIDs) inhibit the cyclooxygenase-1 (COX-1) enzyme, which is responsible for synthesizing cytoprotective prostaglandins in the gut. This leads to a severe drop in mucosal blood flow, a reduction in mucus secretion, and the immediate breakdown of tight junctions, causing epithelial erosion and localized GALT activation within hours [25:1].
What is the role of secretory IgA (sIgA) in food sensitivities?
Secretory IgA acts as a protective shield by binding to dietary proteins in the gut lumen, neutralizing them, and facilitating their non-inflammatory clearance. If mucosal sIgA levels are chronically low, these intact dietary proteins can cross the epithelial barrier, reaching systemic antigen-presenting cells in the lamina propria and triggering inflammatory IgG or IgE-mediated food hypersensitivities [13:1].
Can a high-fiber diet make gut inflammation worse?
Yes, in specific clinical contexts. If a patient suffers from active small intestinal bacterial overgrowth (SIBO) or severe dysbiosis, fermentable fibers will undergo rapid, premature fermentation. This produces organic acids and gases that irritate the gut lining, cause pain, and activate local mast cells, exacerbating inflammatory signaling rather than reducing it [14:1].
¶ Methods & Evidence Grading
Our clinical evaluation prioritizes human randomized controlled trials (RCTs), systematic reviews, and meta-analyses.
- High Certainty: Multiple well-designed human RCTs demonstrating consistent, statistically significant outcomes with direct clinical relevance.
- Moderate Certainty: At least one high-quality RCT or multiple cohort studies showing directional consistency, with minor limitations in sample size or duration.
- Low Certainty: Mechanistic trials, pilot studies, or studies relying heavily on animal models with limited human clinical replication.
¶ References
¶ Update Log
- 2026-07-07: Created the comprehensive clinical guide on the Gut-Immune Axis, featuring cell-mediated signaling, systemic disease axes, and lifespan immunology.
Zhang T, Tang B, Yu Z. (2026). Gut microbiota-immune crosstalk in osteoarthritis: pathogenic mechanisms and emerging therapeutic opportunities. Frontiers in Microbiology. https://pubmed.ncbi.nlm.nih.gov/42404783/ ↩︎ ↩︎ ↩︎ ↩︎
Li Q, Cheng M, Li W. (2026). Oral‑gut axis in systemic disease: A barrier‑metabolism‑immunity three‑dimensional regulatory model (Review). International Journal of Molecular Medicine. https://pubmed.ncbi.nlm.nih.gov/42396665/ ↩︎ ↩︎
Mo M, Chen L, Wang Y. (2026). The gut-lung axis in childhood asthma: from early-life programming to microbiome-informed precision medicine-a narrative review. Frontiers in Immunology. https://pubmed.ncbi.nlm.nih.gov/42099620/ ↩︎
Fan R, Zang Q, Xu Y. (2026). Metagenomic characterization of gut microbiota in rheumatoid arthritis-associated interstitial lung disease: taxonomic shifts and clinical correlations. Frontiers in Immunology. https://pubmed.ncbi.nlm.nih.gov/42367778/ ↩︎ ↩︎ ↩︎
Kamrad S, Gagliani N, Lawley TD. (2026). Gut microbial metabolites for potentiating cancer therapy. Trends in Cancer. https://pubmed.ncbi.nlm.nih.gov/42203551/ ↩︎ ↩︎
Ryan SM, Brayden DJ. (2026). Food-derived molecules as regulators of intestinal tight junctions and barrier function: mechanisms and implications. Frontiers in Drug Delivery. https://pubmed.ncbi.nlm.nih.gov/41939722/ ↩︎ ↩︎ ↩︎ ↩︎
Li S, Liu H. (2026). Gut-bone axis: mechanisms and intervention effects of Chinese botanical drugs in osteoporosis management. Frontiers in Pharmacology. https://pubmed.ncbi.nlm.nih.gov/42389277/ ↩︎
Gu ZY, Cao J, Hu WJ. (2026). [Oral-gut axis: the microbial and immune bridge linking periodontitis to inflammatory bowel disease]. Chinese Journal of Stomatology. https://pubmed.ncbi.nlm.nih.gov/42394565/ ↩︎ ↩︎
Au Yong SJ, Lestari Lee AS, Subramaniyan V. (2026). Gut microbiome modulation in allergic rhinitis: from current evidence to emerging therapies. Frontiers in Allergy. https://pubmed.ncbi.nlm.nih.gov/41993996/ ↩︎ ↩︎
Wang Y, Liu M, Dogra SK. (2026). Effects of an infant formula containing a whey protein concentrate on feeding tolerance and markers of intestinal immune defense in Chinese infants. BMC Nutrition. https://pubmed.ncbi.nlm.nih.gov/42363297/ ↩︎ ↩︎ ↩︎ ↩︎
Wu Y, Shen H, Wang J. (2026). Chinese herbal formulas for Hashimoto's thyroiditis based on the thyroid-gut axis: multitarget synergistic mechanisms and boundaries of evidence. Frontiers in Endocrinology. https://pubmed.ncbi.nlm.nih.gov/42395183/ ↩︎
Shon WJ, Kim KA, Kim JS. (2026). Habitual Ultra-processed Food Intake Is Associated with Gut Dysbiosis and Pro-inflammatory Metabolite Profiles in Korean Patients with IBD. Digestive Diseases and Sciences. https://pubmed.ncbi.nlm.nih.gov/42319657/ ↩︎ ↩︎ ↩︎
Pavačić P, Krpan E, Zeman K. (2026). Gut Microbiota, Metabolic Markers, and Systemic Inflammation in Young Women with Self-Reported Rosacea: An Exploratory Cross-Sectional Study. Journal of Clinical Medicine. https://pubmed.ncbi.nlm.nih.gov/42278997/ ↩︎ ↩︎
Marchitto SA, Abbatecola G, Zeidan RS. (2026). The Gut-Muscle Axis in Sarcopenia: Mechanisms, Evidence Gaps and Translational Challenges. Biomedicines. https://pubmed.ncbi.nlm.nih.gov/42193302/ ↩︎ ↩︎
Wastyk HC, et al. (2021). Gut-microbiota-targeted diets modulate inflammatory signatures and human microbes. Cell. https://pubmed.ncbi.nlm.nih.gov/34256014/ ↩︎ ↩︎ ↩︎ ↩︎
Zhao H, et al. (2026). Multi-target Mechanisms and Clinical Evidence for Ganzaoning Granule in Metabolic Dysfunction-Associated Steatotic Liver Disease: A Narrative Review. Cureus. https://pubmed.ncbi.nlm.nih.gov/4238541/ ↩︎ ↩︎
Caradonna E, Ferrara F, Costantino L. (2026). Clonal Hematopoiesis and Gut Microbiota-Derived TMAO as Candidate Amplifiers of Cardiovascular Inflammation: The CHIDT Hypothesis. Antioxidants. https://pubmed.ncbi.nlm.nih.gov/42352087/ ↩︎ ↩︎
Benjamin J, et al. (2012). Glutamine and whey protein improve intestinal permeability and systemic antigen load in patients with Crohn's disease: a randomized controlled trial. Digestive Diseases and Sciences. https://pubmed.ncbi.nlm.nih.gov/22038507/ ↩︎ ↩︎
Kotani Y, et al. (2014). Oral administration of Lactobacillus rhamnosus GG increases mucosal secretory IgA levels and protective immunity against influenza virus in infant mice. Letters in Applied Microbiology. https://pubmed.ncbi.nlm.nih.gov/24571477/ ↩︎ ↩︎ ↩︎
Zamani B, et al. (2016). Clinical and metabolic response to probiotic supplementation in patients with rheumatoid arthritis: a randomized, double-blind, placebo-controlled trial. International Journal of Rheumatic Diseases. https://pubmed.ncbi.nlm.nih.gov/26200140/ ↩︎
Underwood MA, et al. (2013). Bifidobacterium infantis 35624 in premature infants: a randomized, double-blind, placebo-controlled trial of safety and colonization. Journal of Pediatrics. https://pubmed.ncbi.nlm.nih.gov/23791106/ ↩︎
Marano G, d'Abate C, Ianes I. (2026). The Gut Microbiota in Perimenopausal Anxiety: A Novel Therapeutic Pathway Through Diet. Nutrients. https://pubmed.ncbi.nlm.nih.gov/41829913/ ↩︎
Markle JG, et al. (2013). Sex differences in the gut microbiome drive hormone-dependent regulation of autoimmunity. Science. https://pubmed.ncbi.nlm.nih.gov/23328391/ ↩︎
Franceschi C, et al. (2018). Inflammaging and anti-inflammaging: A systemic view in aging and longevity. Ageing Research Reviews. https://pubmed.ncbi.nlm.nih.gov/29857075/ ↩︎
Mahmood A, et al. (2007). Zinc carnosine, a health food supplement, protects against NSAID-induced increase in gut permeability and mucosal injury in a randomized, double-blind clinical trial. Gut. https://pubmed.ncbi.nlm.nih.gov/16777953/ ↩︎ ↩︎
Langmead L, et al. (2002). Antioxidant effects of herbal therapies used by patients with inflammatory bowel disease: an in vitro study. Alimentary Pharmacology & Therapeutics. https://pubmed.ncbi.nlm.nih.gov/11849138/ ↩︎
Qamar A, et al. (2001). Saccharomyces boulardii stimulates intestinal immunoglobulin A immune response to Clostridium difficile toxin A in mice. Infection and Immunity. https://pubmed.ncbi.nlm.nih.gov/11254634/ ↩︎
Gibson GR, et al. (2017). Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nature Reviews Gastroenterology & Hepatology. https://pubmed.ncbi.nlm.nih.gov/28611480/ ↩︎