¶ Ejercicio
El ejercicio es mucho más que simplemente "estar sano"; es una intervención fisiológica compleja que estimula adaptaciones profundas, influyendo en todo, desde el metabolismo celular hasta la función cognitiva. Comprender sus mecanismos multifacéticos permite establecer protocolos específicos para optimizar la esperanza de vida saludable y la longevidad.

| Indicación | Esperanza de vida saludable y longevidad |
| Acceso | Intervención conductual |
| Frecuencia | Diaria/Semanal (Variada) |
| Perfil de seguridad | Bajo riesgo (si se prescribe adecuadamente) |
| Marcador clave | VO2 máx., Fuerza, Masa magra |
| Costo est. | $0 - $XXX/mes |
La evidencia científica respalda abrumadoramente al ejercicio como un pilar fundamental de la longevidad, principalmente al preservar la masa muscular, mejorar la regulación metabólica a través de las miocinas y mantener una función cardiovascular y neurológica robusta.
¶ De un vistazo
El ejercicio, que abarca tanto el entrenamiento de resistencia como el aeróbico, es una potente intervención de salud. Combate directamente el deterioro relacionado con la edad, mejora la fisiología sistémica y aumenta la calidad de vida. Los beneficios dependen de la dosis, e incluso la actividad moderada produce mejoras significativas en la salud.
Puntos clave:
- Beneficios integrales: El ejercicio impacta casi todos los sistemas fisiológicos, desde la salud cardiovascular hasta la función cognitiva y la regulación metabólica [1][2][3].
- Combate el envejecimiento: Es una estrategia principal para mitigar la sarcopenia (pérdida de masa muscular), preservar la función neuromuscular y reducir la mortalidad por todas las causas [4][5][6].
- Órgano endocrino: El músculo esquelético actúa como un órgano endocrino, secretando miocinas que median los beneficios sistémicos para la salud [3:1][7].
- Seguridad: Generalmente seguro, pero requiere una progresión adecuada y la consideración del estado de salud individual y las contraindicaciones [8].
Para qué se utiliza:
- Objetivos principales: Longevidad, Salud metabólica, Salud cardiovascular, Preservación muscular, Mejora cognitiva, Regulación del estado de ánimo.
- Calidad de la evidencia: Alta (para la salud general y la reducción de la mortalidad).
¶ Tabla resumen de evidencia (resultados en humanos)
| Resultado / Objetivo | Efecto* | Consistencia** | Calidad de la evidencia | Ensayos*** | Notas (población, duración, dosis) |
|---|---|---|---|---|---|
| Mortalidad por todas las causas | Alta | Alta | Metaanálisis de cohortes | Entrenamiento de resistencia: 15-21% menor riesgo [4:1][5:1]; Aeróbico: 13% de reducción por cada aumento de 1 MET [1:1] | |
| Mortalidad cardiovascular | Alta | Alta | Metaanálisis de cohortes | Entrenamiento de resistencia: 19% menor riesgo [5:2] | |
| Mortalidad por cáncer | Moderada | Alta | Metaanálisis de cohortes | Entrenamiento de resistencia: 14% menor riesgo [5:3] | |
| Sarcopenia (pérdida muscular) | Alta | Alta | ECA, estudios longitudinales | El entrenamiento de resistencia aumenta significativamente la masa muscular y la fuerza en adultos mayores [9][10] | |
| Aptitud cardiorrespiratoria (VO2 máx.) | Alta | Alta | Estudios longitudinales | Se correlaciona directamente con la longevidad; las mejoras reducen la mortalidad [2:1][11] | |
| Sensibilidad a la insulina | Alta | Alta | ECA, estudios mecanicistas | Mejora a través de miocinas (ej., IL-6, FGF21) y efectos musculares directos [3:2][12] | |
| Pardeamiento del tejido adiposo | Moderada | Moderada | Mecanicistas, observacionales en humanos | Inducido por miocinas como la irisina y BAIBA [13][14] | |
| Función cognitiva | Moderada | Moderada | Mecanicistas, observacionales, algunos ECA | Mediado por BDNF y otros factores neurotróficos [15] |
*Efecto: El número de flechas (1-3) indica la magnitud. Dirección: ↑ (aumento), ↓ (disminución), = (sin efecto), ? (poco claro). Impacto en la salud: (p) = positivo para la salud, (n) = negativo para la salud, (x) = impacto neutral/desconocido. Ejemplos: ↓↓↓ (p) = gran disminución, positivo; ↑ (n) = pequeño aumento, negativo; = (x) = sin efecto; ? = poco claro.
*Codificación del renderizador compacto (preferido al usar etiquetas personalizadas): <effect e="[dir][mag][impact]"></effect> donde dir = u|d|e|q, mag = 0|1|2|3, impact = p|n|x. Ejemplos: ↓↓ (p) -> <effect e="d2p"></effect>, = (x) -> <effect e="e0x"></effect>, ? -> <effect e="q0x"></effect>
**Consistencia: Baja (resultados contradictorios), Moderada (mixta pero con tendencia hacia un lado), Alta (la mayoría de los ensayos coinciden)
***Ensayos: Número de ECA o ensayos totales que informan este resultado (muestra la profundidad de la evidencia de un vistazo)
OBLIGATORIO: DEBE incluir una clave de cita (ej. [^1]) en la columna "Notas" para cada fila. Si afirma un resultado, debe enlazar el Metaanálisis específico o ECA clave que lo demuestre.
¶ Mecanismos de acción
El ejercicio produce sus amplios beneficios a través de varios mecanismos fisiológicos interconectados:
| Mecanismo | Efecto principal | Resultados clave | Evidencia |
|---|---|---|---|
| Preservación de la masa muscular y la fuerza | Ralentiza la sarcopenia (pérdida muscular relacionada con la edad) | Mantiene la movilidad, reduce el riesgo de discapacidad y mortalidad | Cohortes observacionales; ECA con ganancias de hipertrofia/fuerza [6:1][9:1][10:1] |
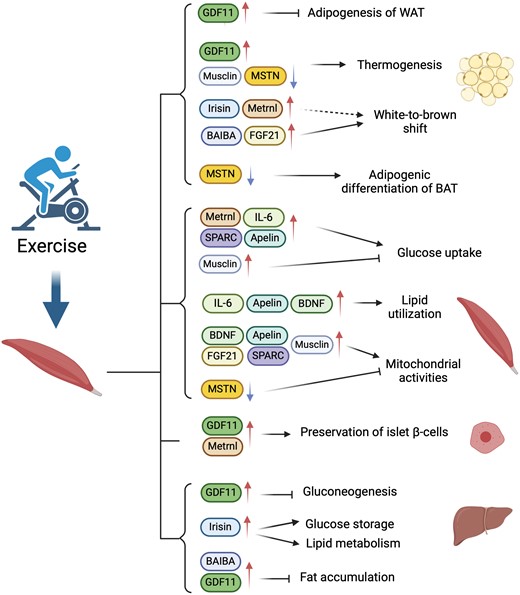
| Señalización endocrina (miocinas) | IL‑6, irisina, miostatina, IL‑15, BDNF, FGF21, BAIBA, METRNL, mionectina, SPARC, decorina, apelina | Mejora la sensibilidad a la insulina, la oxidación de lípidos, el pardeamiento adiposo, la resolución de la inflamación, la reparación de tejidos | Mecanicistas, ejercicio agudo en humanos, ECA/estudios controlados [3:3][7:1][13:1][15:1] |
| Preservación del tipo de fibra y neuromuscular | Contrarresta la atrofia/denervación preferencial de las fibras tipo II | Mantiene la potencia, reduce el riesgo de caídas, apoya la independencia funcional | Histología, fisiología longitudinal, ensayos de intervención [6:2][9:2][10:2] |
| Adaptaciones cardiorrespiratorias | Mejora el gasto cardíaco, la función vascular, la densidad mitocondrial | Mejora el VO2 máx., reduce el riesgo de enfermedades cardiovasculares | Metaanálisis, estudios fisiológicos [1:2][2:2] |
Una ilustración científica que demuestra cómo el ejercicio desencadena la liberación de miocinas del músculo esquelético, ilustrando la compleja respuesta metabólica del cuerpo a la actividad física.
¶ 1) Masa muscular y autonomía: combatiendo la sarcopenia
La sarcopenia, la pérdida progresiva de masa muscular esquelética, fuerza y función con el envejecimiento, es un importante predictor de discapacidad, caídas y mortalidad [3:4][6:3]. Los análisis cuantitativos muestran que la fuerza disminuye más rápido que la masa muscular, lo que indica cambios en la calidad muscular [16].
El entrenamiento de resistencia es altamente efectivo para contrarrestar la sarcopenia:
- Reversibilidad: Los estudios en adultos mayores y muy mayores (edad media ~87 años) demuestran que el entrenamiento de resistencia de alta intensidad puede producir ganancias clínicamente significativas en la fuerza y en el área de sección transversal (CSA) de las fibras, incluso en individuos frágiles [9:3][10:3].
- Ganancias funcionales: Estas ganancias de fuerza se traducen en una mejor movilidad y un menor riesgo de caídas, lo cual es crucial para mantener la autonomía y la calidad de vida en la edad avanzada [10:4].
¶ 2) Los músculos como órganos endocrinos: el papel de las mioquinas
El músculo esquelético secreta activamente diversas citoquinas y péptidos, conocidos colectivamente como mioquinas, en la circulación durante y después de la contracción [3:5][7:2]. Estas mioquinas actúan de manera autocrina, paracrina y endocrina para regular el metabolismo de todo el cuerpo, la inflamación, la reparación de tejidos y la función cerebral.
¶ Principales mioquinas (seleccionadas) y sus acciones principales
| Mioquina (acrónimo expandido una vez) | Acciones principales | Contexto típico | Evidencia |
|---|---|---|---|
| Interleucina‑6 (IL‑6) | Aumenta la captación de glucosa y la oxidación de ácidos grasos en el músculo; lipólisis; señalización antiinflamatoria aguda durante el ejercicio | Ejercicio agudo → aumento transitorio | Estudios mecanicistas y de ejercicio en humanos [3:6][17] |
| Irisina (escindida de FNDC5) | Induce el pardeamiento del tejido adiposo blanco; aumenta la termogénesis y el gasto energético | Ejercicio de resistencia/fuerza | Descubrimiento en Nature; datos de observación/intervención en humanos [13:2][18] |
| Miostatina (GDF‑8) | Regulador negativo del crecimiento muscular; inhibición → hipertrofia | Regulación basal; se reduce con el entrenamiento | Genética/mecanicista; relevancia traslacional [19] |
| Interleucina‑15 (IL‑15) | Apoya el anabolismo muscular; asociada con una menor adiposidad | Sensible al ejercicio | Estudios biológicos/en humanos [20] |
| Factor neurotrófico derivado del cerebro (BDNF) | Aumenta la oxidación de grasas a través de AMPK; eje músculo-cerebro (beneficios neurotróficos y cognitivos) | Contracción muscular; ejercicio de resistencia | Trabajo mecanicista y en células humanas [15:2][20:1] |
| Factor de crecimiento de fibroblastos‑21 (FGF21) | Mejora la sensibilidad a la insulina; regula el metabolismo de glucosa/lípidos | Ejercicio agudo y crónico; estrés metabólico | Ejercicio/fisiología en humanos/ratones [12:1][21] |
| Ácido β‑aminoisobutírico (BAIBA) | Promueve la β‑oxidación hepática; induce el pardeamiento de la grasa blanca | Metabolito inducido por el ejercicio | Asociaciones en humanos; datos mecanicistas [14:1] |
| Similar a la meteorina (METRNL) | Mejora la termogénesis de la grasa beige; interacciones inmunitario-adiposas | Frío/ejercicio | Trabajo en células y animales con relevancia traslacional [22] |
| Mionectina (CTRP15) | Aumenta la captación de ácidos grasos; activa mTOR; suprime la autofagia hepática | Ejercicio/contracción muscular | Estudios mecanicistas e in vivo [23][24] |
| Proteína ácida rica en cisteína secretada (SPARC) | Efectos metabólicos; señalización antitumorigénica en el colon vinculada al ejercicio | Ejercicio regular | Datos mecanicistas en animales y tejidos humanos [25] |
| Decorina | Se une/inhibe a la miostatina; contribuye a la hipertrofia | Ejercicio de fuerza | Estudios mecanicistas en humanos/animales [26] |
| Apelina | Mejora la función/regeneración muscular; efectos metabólicos favorables | Entrenamiento aeróbico; músculo envejecido | Datos de cohortes/intervención en humanos [25:1] |
Notas: Las elevaciones agudas de IL‑6 durante el ejercicio difieren de la inflamación crónica de bajo grado; el contexto (tiempo/tejido) determina el efecto neto [3:7][17:1]. La evidencia abarca estudios mecanicistas, de ejercicio agudo en humanos y de entrenamiento controlado; no todas las mioquinas tienen ensayos de resultados clínicos definitivos todavía [7:3][20:2].
¶ 3) La pérdida de fibras tipo II (contracción rápida) con el envejecimiento
El músculo esquelético humano comprende dos tipos principales de fibras: Tipo I (contracción lenta), que son resistentes a la fatiga y se utilizan para la resistencia, y Tipo II (contracción rápida), que generan gran fuerza y potencia pero se fatigan rápidamente. El envejecimiento no afecta a estas fibras por igual.

Esquema que ilustra la pérdida preferencial de fibras musculares tipo II (contracción rápida) en comparación con las fibras tipo I (contracción lenta) a lo largo de la vida humana. Mientras que las fibras tipo I permanecen relativamente estables, las fibras tipo II sufren una atrofia y pérdida significativas a partir de la mediana edad. Modelado a partir de los datos de Lexell et al. [6:4]
¶ Atrofia y pérdida preferencial
La pérdida de músculo relacionada con la edad es predominantemente una pérdida de fibras tipo II:
- Atrofia selectiva: Las fibras tipo II se encogen significativamente en tamaño (área de sección transversal), reduciéndose en un 25–40% o más entre los 20 y los 80 años [6:5][27]. Las fibras tipo I, por el contrario, generalmente mantienen su tamaño.
- Pérdida de unidades motoras: Las motoneuronas alfa que inervan las fibras tipo II son propensas a la apoptosis (muerte celular). A medida que estas neuronas mueren, las fibras musculares asociadas pierden su conexión con el sistema nervioso [6:6].
¶ El mecanismo de "agrupación"
Cuando una fibra de Tipo II se denerva, pide ayuda. A menudo, una neurona motora de Tipo I cercana desarrollará un nervio colateral para reinervar la fibra "huérfana". Esto salva a la fibra de morir, pero tiene un costo: la fibra se convierte de Tipo II a Tipo I [9:4][27:1].
- Resultado: El músculo se vuelve más lento y menos potente.
- Arquitectura agrupada: En lugar del patrón saludable de "tablero de ajedrez" de tipos de fibras mixtas que se observa en la juventud, el músculo envejecido muestra una "agrupación" de tipos de fibras, lo que indica este historial de denervación y reinervación [6:7].
¶ Consecuencias clínicas: Potencia vs. Fuerza
Debido a que las fibras de Tipo II impulsan el movimiento de alta velocidad, la potencia muscular (la capacidad de ejercer fuerza rápidamente) disminuye mucho más rápido que la fuerza pura [27:2]. Esto tiene implicaciones críticas para la independencia:
- Riesgo de caídas: Recuperarse de un tropiezo requiere un paso explosivo en una fracción de segundo (actividad de Tipo II).
- Movilidad: Levantarse de una silla o subir escaleras requiere potencia.
- Solución: El entrenamiento de resistencia, particularmente con la intención de mover el peso rápidamente (entrenamiento de potencia), puede hipertrofiar las fibras de Tipo II restantes y mejorar las tasas de disparo de las unidades motoras, revirtiendo parcialmente este declive [9:5][10:5].
¶ 4) Adaptaciones cardiorrespiratorias
El ejercicio aeróbico regular mejora significativamente la aptitud cardiorrespiratoria, medida por el VO2 max. Una mayor aptitud cardiorrespiratoria está fuerte e inversamente asociada con la mortalidad por todas las causas, los eventos cardiovasculares y otras enfermedades no transmisibles importantes [1:3][2:3].
- Eficiencia cardíaca: El entrenamiento con ejercicios conduce a un aumento del volumen sistólico, una reducción de la frecuencia cardíaca en reposo y una mejora de la contractilidad miocárdica, mejorando la eficiencia de bombeo del corazón.
- Salud vascular: Promueve la función endotelial, mejora la elasticidad arterial y reduce la resistencia vascular sistémica, disminuyendo así la presión arterial y reduciendo el riesgo de aterosclerosis.
- Biogénesis mitocondrial: Tanto el entrenamiento aeróbico como el de resistencia estimulan el crecimiento de nuevas mitocondrias dentro de las células musculares, mejorando la producción de energía y la flexibilidad metabólica [3:8].
¶ Protocolos prácticos e integración
Un régimen de ejercicio integral debe incluir una combinación de entrenamiento de resistencia, aeróbico, de flexibilidad y de equilibrio. Para obtener el máximo beneficio de longevidad sin sobreentrenamiento, estas modalidades deben estructurarse dentro de un plan organizado. Consulte la guía detallada sobre Bloques de entrenamiento y periodización para aprender cómo secuenciar macrociclos, mesociclos y microciclos de manera efectiva.
¶ Entrenamiento de resistencia
- Frecuencia: 2-3 veces por semana en días no consecutivos.
- Intensidad: 70-85% de una repetición máxima (1-RM) para 8-12 repeticiones por serie, o RPE (Índice de Esfuerzo Percibido) 7-9.
- Volumen: 2-4 series por grupo muscular principal.
- Tipo: Incorpore movimientos compuestos (sentadillas, pesos muertos, press, remos) que se dirijan a múltiples grupos musculares.
- Progresión: Aumente gradualmente el peso, las repeticiones o las series con el tiempo para garantizar una sobrecarga progresiva.
- Enfoque en la potencia: Integre periódicamente movimientos con la intención de moverse rápidamente para apuntar a las fibras de Tipo II.
¶ Entrenamiento aeróbico
- Frecuencia: 3-5 veces por semana.
- Intensidad: Moderada (se puede hablar pero no cantar) durante 150-300 minutos por semana, o vigorosa (difícil hablar) durante 75-150 minutos por semana.
- Tipo: Caminar, trotar, andar en bicicleta, nadar, bailar u otras actividades que eleven la frecuencia cardíaca.
- HIIT: El entrenamiento en intervalos de alta intensidad se puede incorporar 1-2 veces por semana para mejorar la aptitud cardiorrespiratoria y las adaptaciones metabólicas.
¶ Flexibilidad y equilibrio
- Frecuencia: Diaria o 2-3 veces por semana.
- Tipo: Estiramientos, yoga, tai chi o ejercicios específicos de equilibrio.
- Beneficios: Mejora el rango de movimiento, reduce el riesgo de lesiones y mantiene la movilidad.
¶ Seguimiento de métricas
- Fuerza: Evalúe periódicamente la 1-RM o las repeticiones máximas para los levantamientos clave.
- Aptitud cardiorrespiratoria: Estime o mida directamente el VO2 max (por ejemplo, utilizando monitores de actividad física o pruebas de laboratorio).
- Masa muscular magra: Supervise los cambios mediante escáneres DEXA, análisis de impedancia bioeléctrica (BIA) o mediciones regulares de circunferencia.
- Resultados funcionales: Realice un seguimiento de las mejoras en las actividades de la vida diaria, la velocidad de la marcha y las pruebas de equilibrio.
¶ Seguridad y contraindicaciones
Aunque el ejercicio es generalmente seguro y muy beneficioso, ciertas condiciones requieren precaución o autorización médica [8:1]:
¶ Precauciones generales
- Progresión gradual: Comience con baja intensidad y volumen, especialmente en personas sedentarias, y aumente gradualmente a lo largo de semanas o meses.
- Escuche a su cuerpo: El dolor es una señal para detener o modificar el ejercicio. Diferencie entre el dolor muscular y el dolor articular o de ligamentos.
- Hidratación y nutrición: Asegure una ingesta adecuada de líquidos y una sincronización apropiada de los macronutrientes para favorecer el rendimiento y la recuperación.
¶ Contraindicaciones absolutas (Evite el ejercicio hasta que un médico lo autorice)
- Angina inestable
- Infarto de miocardio reciente (en los últimos 2 días)
- Arritmia cardíaca no controlada que causa síntomas o compromiso hemodinámico
- Endocarditis, miocarditis o pericarditis aguda
- Estenosis aórtica severa sintomática
- Insuficiencia cardíaca descompensada
- Embolia pulmonar aguda o infarto pulmonar
- Tromboflebitis aguda
- Aneurisma disecante
- Infección sistémica aguda con fiebre, dolores corporales o ganglios linfáticos inflamados
¶ Contraindicaciones relativas (El ejercicio puede realizarse con precaución o bajo supervisión)
- Estenosis obstructiva conocida de la arteria coronaria principal izquierda
- Estenosis aórtica de moderada a severa con estado de síntomas incierto
- Taquiarritmias con frecuencias ventriculares no controladas
- Bloqueo cardíaco adquirido avanzado o completo
- Accidente cerebrovascular reciente o ataque isquémico transitorio
- Deterioro mental con cooperación limitada
- Hipertensión en reposo (sistólica >200 mmHg o diastólica >110 mmHg)
- Condiciones médicas no corregidas, como anemia significativa, desequilibrio electrolítico, hipertiroidismo
¶ Preguntas frecuentes
¿Cuánto ejercicio necesito para ver beneficios en la longevidad?
Para la salud general y la longevidad, las principales organizaciones de salud recomiendan al menos 150 minutos de actividad aeróbica de intensidad moderada o 75 minutos de actividad aeróbica de intensidad vigorosa por semana, combinados con actividades de fortalecimiento muscular 2 o más días a la semana. Sin embargo, la evidencia sugiere una relación dosis-respuesta, con beneficios adicionales observados a volúmenes más altos, particularmente para la aptitud cardiorrespiratoria [5:4][1:4].
¿Puedo hacer demasiado ejercicio?
Si bien los beneficios del ejercicio son sustanciales, el entrenamiento excesivo sin una recuperación adecuada puede provocar el síndrome de sobreentrenamiento, un mayor riesgo de lesiones, desequilibrios hormonales y una función inmunológica deteriorada. La tolerancia individual varía, pero equilibrar el estrés del entrenamiento con la recuperación es crucial [7:4].
¿Es mejor el entrenamiento de resistencia o el ejercicio aeróbico para la longevidad?
Ambos son fundamentales. El entrenamiento de resistencia es esencial para preservar la masa y la fuerza muscular, combatir la sarcopenia y mejorar la salud metabólica [9:6]. El ejercicio aeróbico mejora la capacidad cardiorrespiratoria, la función endotelial y la salud mitocondrial [1:5]. Combinar ambos tipos de ejercicio proporciona una reducción superior de la mortalidad por todas las causas en comparación con cualquiera de ellos por separado [28].
¿Cómo puedo hacer del ejercicio un hábito constante?
Empieza poco a poco y aumenta gradualmente. Encuentra actividades que disfrutes, establece objetivos realistas e integra el ejercicio en tu rutina diaria. Contar con compañeros ante los que rendir cuentas, hacer un seguimiento del progreso y variar tus entrenamientos también puede ayudar a mantener la constancia [8:2].
¶ Referencias
Kodama S, Saito K, Tanaka S, et al. Cardiorespiratory fitness as a quantitative predictor of all-cause mortality and cardiovascular events in healthy men and women: a meta-analysis. JAMA. 2009;301(19):2024-2035. https://pubmed.ncbi.nlm.nih.gov/19454641/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Mandsager K, et al. Association of Cardiorespiratory Fitness With Long-term Mortality Among Adults Undergoing Exercise Treadmill Testing. JAMA Network Open. 2018;1(6):e183605. https://jamanetwork.com/journals/jamanetworkopen/fullarticle/2707428 ↩︎ ↩︎ ↩︎ ↩︎
Pedersen BK, Febbraio MA. Muscle as an endocrine organ: focus on muscle‑derived interleukin‑6. Physiol Rev. 2008;88(4):1379‑1406. https://journals.physiology.org/doi/full/10.1152/physrev.90100.2007 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Saeidifard F, Medina-Inojosa JR, Welton M, et al. The association of resistance training with mortality: A systematic review and meta-analysis. Eur J Prev Cardiol. 2019;26(15):1647-1662. https://pubmed.ncbi.nlm.nih.gov/31104484/ ↩︎ ↩︎
Shailendra P, et al. Resistance Training and Mortality Risk: A Systematic Review and Meta-Analysis. Am J Prev Med. 2022;63(4):603-611. https://pubmed.ncbi.nlm.nih.gov/35599175/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Lexell J, Taylor CC, Sjöström M. Ageing atrophy: number/size/proportion of fiber types in vastus lateralis (15–83 y). J Neurol Sci. 1988;84:275‑294. https://www.sciencedirect.com/science/article/abs/pii/0022510X88901245 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Severinsen MCK, Pedersen BK. Muscle–organ crosstalk: the emerging roles of myokines. Endocr Rev. 2020;41(4):594‑609. https://pubmed.ncbi.nlm.nih.gov/32374815/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Riebe D, et al. Resistance Exercise Training in Individuals With and Without Cardiovascular Disease: 2023 Update. Circulation. 2023;147(14):1128-1151. https://www.ahajournals.org/doi/10.1161/CIR.0000000000001189 ↩︎ ↩︎ ↩︎
Frontera WR, Meredith CN, O’Reilly KP, Knuttgen HG, Evans WJ. Strength conditioning in older men: hypertrophy and function. J Appl Physiol. 1988;64(3):1038‑1044. https://journals.physiology.org/doi/abs/10.1152/jappl.1988.64.3.1038 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Fiatarone MA, O’Neill EF, Ryan ND, et al. Exercise training in very elderly people (RCT). N Engl J Med. 1994;330(25):1769‑1775. https://www.nejm.org/doi/full/10.1056/NEJM199406233302501 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Laukkanen JA, et al. Long-term Change in Cardiorespiratory Fitness and All-Cause Mortality: A Population-Based Follow-up Study. Mayo Clin Proc. 2016;91(8):1021-1028. https://pubmed.ncbi.nlm.nih.gov/27444976/ ↩︎
Izumiya Y, et al. FGF21 is an Akt‑regulated myokine. FEBS Lett. 2008;582(27):3805‑3810. https://febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2008.10.021 ↩︎ ↩︎
Boström P, et al. A PGC1‑α‑dependent myokine (irisin) that drives browning of white fat. Nature. 2012;481:463‑468. https://www.nature.com/articles/nature10777 ↩︎ ↩︎ ↩︎
Roberts LD, et al. β‑Aminoisobutyric acid induces browning of white fat and hepatic β‑oxidation. Cell Metab. 2014;19(1):96‑108. https://www.cell.com/cell-metabolism/fulltext/S1550-4131(13)00478-0 ↩︎ ↩︎
Wrann CD, et al. Exercise induces hippocampal BDNF via a PGC‑1α/FNDC5 pathway. Cell Metab. 2013;18(5):649‑659. https://www.cell.com/cell-metabolism/fulltext/S1550-4131(13)00372-3 ↩︎ ↩︎ ↩︎
Goodpaster BH, et al. Loss of strength, mass, and quality in older adults (Health ABC). J Gerontol A. 2006;61(10):1059‑1064. https://academic.oup.com/biomedgerontology/article/61/10/1059/545851 ↩︎
Akerström T, et al. Exercise induces IL‑6 release from human skeletal muscle; role in lipid metabolism. Am J Physiol Endocrinol Metab. 2005;288:E733‑E740. https://journals.physiology.org/doi/full/10.1152/ajpendo.00340.2004 ↩︎ ↩︎
Huh JY, et al. Irisin stimulates muscle growth‑related genes and regulates adipocyte metabolism in humans. Int J Obes (Lond). 2014;38:1538‑1544. https://www.nature.com/articles/ijo201442 ↩︎
McPherron AC, Lawler AM, Lee SJ. Regulation of skeletal muscle mass by a new TGF‑β family member (myostatin). Nature. 1997;387:83‑90. https://www.nature.com/articles/387083a0 ↩︎
Green DJ, et al. The Potential Role of Contraction‑Induced Myokines in Metabolic Regulation. Front Endocrinol (Lausanne). 2017;8:97. https://www.frontiersin.org/articles/10.3389/fendo.2017.00097/full ↩︎ ↩︎ ↩︎
Kim KH, Lee MS, et al. Acute exercise induces FGF21 in mice and humans. PLoS One. 2013;8(5):e63517. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0063517 ↩︎
Rao RR, et al. Meteorin‑like regulates beige fat thermogenesis. Cell. 2014;157(6):1279‑1291. https://www.cell.com/fulltext/S0092-8674(14)00578-3 ↩︎
Seldin MM, et al. Myonectin (CTRP15) links skeletal muscle to systemic lipid homeostasis. J Biol Chem. 2012;287(15):11968‑11980. https://www.jbc.org/article/S0021-9258(20)50094-2/fulltext ↩︎
Seldin MM, et al. Skeletal muscle‑derived myonectin activates mTOR to suppress hepatic autophagy. J Biol Chem. 2013;288(50):36073‑36082. https://www.jbc.org/article/S0021-9258(20)50092-3/fulltext ↩︎
Aoi W, Naito Y, Takagi T, et al. SPARC and exercise‑linked suppression of colon tumorigenesis. Gut. 2013;62(6):882‑889. https://gut.bmj.com/content/62/6/882.long ↩︎ ↩︎
Kanzleiter T, Rath M, Görgens SW, et al. The myokine decorin is regulated by contraction and involved in hypertrophy. Biochem Biophys Res Commun. 2014;450(2):1089‑1094. https://www.sciencedirect.com/science/article/pii/S0006291X14011089 ↩︎
Verdijk LB, et al. Skeletal muscle hypertrophy following resistance training is accompanied by a fiber type-specific increase in satellite cell content in elderly men. J Gerontol A Biol Sci Med Sci. 2009;64(3):332-339. https://academic.oup.com/biomedgerontology/article/64A/3/332/625396 ↩︎ ↩︎ ↩︎
Momma H, Kawakami R, Honda T, et al. Muscle-strengthening activities are associated with lower risk and mortality in major non-communicable diseases: a systematic review and meta-analysis of cohort studies. Br J Sports Med. 2022;56(13):755-763. https://bjsm.bmj.com/content/56/13/755 ↩︎