¶ MicroARN
Los microARN (miARN) representan una clase altamente conservada de moléculas de ARN endógenas, cortas (típicamente de 19 a 25 nucleótidos) y no codificantes que ejercen un control postranscripcional sobre la expresión génica. Al hibridarse con secuencias complementarias dentro de las regiones 3' no traducidas (UTR) de los ARN mensajeros (ARNm) diana, los miARN inician la represión traduccional o la escisión enzimática, "afinando" eficazmente el proteoma celular.
En el contexto de la biogerontología y la medicina clínica de la longevidad, los microARN desempeñan un papel dual crítico. En primer lugar, actúan como biomarcadores altamente estables y accesibles en biofluidos ("senomiRs" e "inflamma-miRs") que reflejan las trayectorias del envejecimiento sistémico, la disfunción específica de órganos y la carga de células senescentes. En segundo lugar, funcionan como dianas e intervenciones terapéuticas, donde se despliegan imitadores de miARN sintéticos o inhibidores antisentido (antagomirs) para reprogramar vías patológicas y restaurar la homeostasis celular juvenil.
| Indicación | Investigación / Fuera de indicación (Vías de longevidad, enfermedades relacionadas con la edad) |
| Acceso | En investigación / Solo para investigación / Ensayos de fase II y III |
| Esquema de dosificación | Dependiente de la administración (p. ej., subcutánea semanal, intradérmica local o moléculas pequeñas orales diarias) |
| Perfil de seguridad | Moderado (Depende de la especificidad del vector, la inmunogenicidad y los riesgos fuera del objetivo) |
| Marcador clave | Perfil de miARN circulante, niveles de transcripción de genes diana, paneles inflamatorios sistémicos |
| Costo est. | Alto (Terapias de grado clínico en investigación) |
¶ De un vistazo
Los microARN funcionan como reguladores postranscripcionales, sirviendo como nodos fundamentales en las redes de longevidad. Su capacidad para modular simultáneamente múltiples genes posteriores los convierte en dianas clínicas excepcionalmente poderosas, aunque muy complejas.
¶ Puntos clave
- Regulación sistémica: Un solo microARN puede coordinar redes genéticas enteras, influyendo en las marcas del envejecimiento como la senescencia celular, la inflamación crónica y la regeneración de tejidos [1].
- Precisión diagnóstica: Los miARN extracelulares empaquetados en exosomas son excepcionalmente estables en circulación, proporcionando lecturas diagnósticas no invasivas de la edad biológica y la carga de células de senescencia celular [2][3].
- Cuellos de botella en la administración: La traducción clínica de las terapias con miARN está fuertemente limitada por la seguridad del vector de administración, ya que las nanopartículas lipídicas (LNPs) de primera generación pueden desencadenar reacciones inmunitarias graves, y la administración viral no controlada puede provocar un crecimiento tisular aberrante [4][5].
¶ Para qué se utiliza
- Objetivos principales: Eliminación sistémica de la senescencia, regeneración del tejido miocárdico y cutáneo, supresión de la inflamación vascular crónica y perfil diagnóstico del envejecimiento biológico.
- Calidad de la evidencia: Alta para intervenciones localizadas (fibrosis, dianas virales) y biomarcadores observacionales; de moderada a baja para terapias sistémicas de longevidad en desarrollo clínico en etapa temprana.
¶ ¿Qué es?
Los microARN son moléculas de ARN endógenas que actúan como los "atenuadores" maestros del genoma de los mamíferos. Descubiertos por ser fundamentales en la sincronización del desarrollo y la diferenciación celular, no codifican proteínas, sino que gobiernan la estabilidad y la traducción de los ARNm codificantes de proteínas [1:1].
A diferencia de los oligonucleótidos antisentido (ASOs) clásicos que se dirigen a un único transcrito específico, un solo miRNA normalmente posee una "secuencia semilla" (nucleótidos 2–8) que coincide con elementos de consenso en docenas o incluso cientos de mRNAs distintos. Esta capacidad de múltiples objetivos permite a los miRNAs actuar como reguladores a nivel de sistemas, coordinando respuestas fisiológicas complejas como la defensa contra el estrés celular, la remodelación metabólica y las cascadas de señalización inflamatoria.
Modalidades Terapéuticas Principales
- miRNA Mimics: Moléculas de ARN bicatenario sintetizadas químicamente, diseñadas para reemplazar o reforzar un miRNA protector específico que ha disminuido durante el envejecimiento biológico (por ejemplo, miR-29 para la antifibrosis [6] o miR-34a en la supresión de oncogenes [4:1]).
- Antagomirs (Anti-miRs): Oligonucleótidos monocatenarios modificados químicamente, diseñados para unirse con alta afinidad a miRNAs endógenos que promueven enfermedades o son pro-senescentes, neutralizándolos y evitando que silencien los mRNAs diana protectores (por ejemplo, miravirsen dirigido al miR-122 hepático [7] o cobomarsen dirigido al miR-155 proinflamatorio [8]).
- Moduladores Indirectos: Terapias de moléculas pequeñas que regulan al alza o a la baja de manera selectiva la transcripción o el procesamiento de miRNAs específicos en los tejidos diana, como el obefazimod, que mejora selectivamente el regulador antiinflamatorio miR-124 [9].
¶ Beneficios Principales
Dirigirse a los microARN ofrece ventajas incomparables en el diseño terapéutico debido a su capacidad para regular redes celulares integradas en lugar de receptores aislados individuales.
¶ Perfilado Sistémico de la Firma de Longevidad
Los estudios epidemiológicos y de cohortes han demostrado que los centenarios y las personas longevas albergan perfiles distintos de microARN circulantes. Por ejemplo, los nonagenarios y centenarios de las cohortes de Okinawa exhiben una "firma de longevidad" protectora caracterizada por la regulación al alza sostenida de miRNAs antiinflamatorios (como miR-181a-5p) y la regulación a la baja de miRNAs proinflamatorios y pro-senescentes (como miR-21-5p) [10]. La medición de estos paneles permite a los médicos trazar las trayectorias de los pacientes a lo largo de vías de envejecimiento biológico saludables frente a no saludables [11][3:1].
¶ Reversión de la Senescencia y la Fibrosis
A medida que los tejidos envejecen, la acumulación de células senescentes y el depósito progresivo de matriz extracelular colagenosa conducen a la fibrosis parenquimatosa y al endurecimiento de los órganos. Las terapias basadas en miRNA han demostrado la capacidad de revertir estos cambios en modelos clínicos:
- Remodelación Tisular: La administración intradérmica de miRNA mimics de miR-29 (remlarsen) reprime directamente la expresión de colágeno-1, colágeno-3 y otras proteínas de la matriz extracelular, restaurando la elasticidad juvenil del tejido y previniendo la formación patológica de cicatrices [6:1].
- Diagnóstico con SenomiR: Los paneles de 22 miRNAs circulantes (senomiRs) pueden cuantificar con precisión la carga sistémica de células en senescencia celular, proporcionando una lectura molecular directa de la eficacia de los fármacos senolíticos [2:1].
¶ Reprogramación antiinflamatoria (Inflamma-miRs)
La inflamación crónica, de bajo grado y estéril ("inflammaging" o inflamación asociada al envejecimiento) está mediada por la regulación al alza progresiva de los miARN proinflamatorios. Los antagomiRs sistémicos pueden detener esta cascada. Por ejemplo, la neutralización de miR-155 mediante cobomarsen suprime la activación patogénica de las células T y regula a la baja las citoquinas proinflamatorias sistémicas, presentando una vía terapéutica para la inflamación crónica [8:1]. De manera similar, la regulación al alza del "freno" celular miR-124 utilizando la molécula pequeña obefazimod impulsa reducciones significativas y duraderas en las citoquinas inflamatorias (por ejemplo, IL-1β, IL-6, TNF-α), lo que conduce a una remisión clínica y endoscópica profunda en ensayos clínicos [9:1].
¶ Tabla resumen de evidencia (Resultados en humanos)
| Resultado / Objetivo | Efecto* | Consistencia** | Calidad de la evidencia | Ensayos*** | Notas (población, duración, dosis) |
|---|---|---|---|---|---|
| Supresión viral de la hepatitis C | Alta | Alta | 1 ECA de fase 2a | La dosificación subcutánea semanal de miravirsen (anti-miR-122) durante 29 días redujo de manera significativa y duradera la carga viral en pacientes con VHC crónico [7:1]. | |
| Remisión de la colitis ulcerosa | Alta | Alta | 1 ECA de fase 2b, Fase 3 | La dosificación oral una vez al día de obefazimod (molécula pequeña ABX464 reguladora al alza de miR-124) redujo significativamente la puntuación de Mayo modificada e indujo la curación de la mucosa a las 8 semanas [9:2]. | |
| Supresión de fibrosis cutánea y cicatrices | Alta | Alta | 1 ECA de fase 1 | Las inyecciones intradérmicas de remlarsen (mimético de miR-29) en sitios de heridas por escisión regulan a la baja de forma segura el colágeno y la MEC [6:2]. | |
| Estabilización del linfoma de células T | Moderada | Moderada | 1 ensayo de fase 1 | El cobomarsen (antagomiR LNA anti-miR-155) administrado sistémicamente condujo a la interacción con el objetivo y a la estabilización de la enfermedad [8:2]. | |
| Angiogénesis y reparación de tejidos | Alta | Moderada | 1 ensayo de fase 1 | La inyección intradérmica de MRG-110 (ASO LNA anti-miR-92a) aceleró la perfusión, la expresión de CD49e y el crecimiento de los vasos sanguíneos [12][13]. | |
| Estabilización de tumores (Mesotelioma) | Moderada | Moderada | 1 ensayo de fase 1 | TargomiRs (mimético de miR-16 a través de minicélulas dirigidas a EGFR) produjo una enfermedad estable en el 68% de los pacientes con mesotelioma [14]. | |
| Supresión de tumores sólidos (LNP sistémico) | Baja | Baja | 1 ensayo de fase 1 | El ensayo de MRX34 (mimético de miR-34a a través de LNP) se detuvo debido a eventos adversos graves mediados por el sistema inmunitario y muertes [4:2]. | |
| Predicción del riesgo de mortalidad por todas las causas | Alta | Moderada | Estudio de cohorte | Los modelos de edad biológica basados en miARN (mirAge, mirMort, mirFI) predicen el deterioro de la salud autoinformado, la fragilidad y la mortalidad por todas las causas a 10 años [3:2]. | |
| Biomarcadores de longevidad extrema | Alta | Moderada | Observacional | Los perfiles séricos de centenarios muestran una regulación al alza protectora de miR-181a-5p y una regulación a la baja del miR-21-5p inflamatorio [10:1]. | |
| Diagnóstico de la carga de senescencia sistémica | Moderada | Moderada | Estudio de cohorte | Un panel de 22 senomiRs circulantes (incluyendo miR-31-5p, miR-19a-3p) aumenta de manera estable con la edad y se correlaciona con la acumulación de células senescentes en los tejidos [2:2][15]. |
- *Efecto: La codificación del renderizador compacto representa la dirección y la magnitud del resultado terapéutico o del biomarcador.
- **Consistencia: Baja (resultados clínicos contradictorios), Moderada (mixta pero con tendencia positiva), Alta (la mayoría de los ensayos y estudios concuerdan).
- ***Ensayos: Número de ECA (ensayos controlados aleatorizados), ensayos clínicos o cohortes prospectivas que informan el criterio de valoración clínico.
¶ Mecanismos
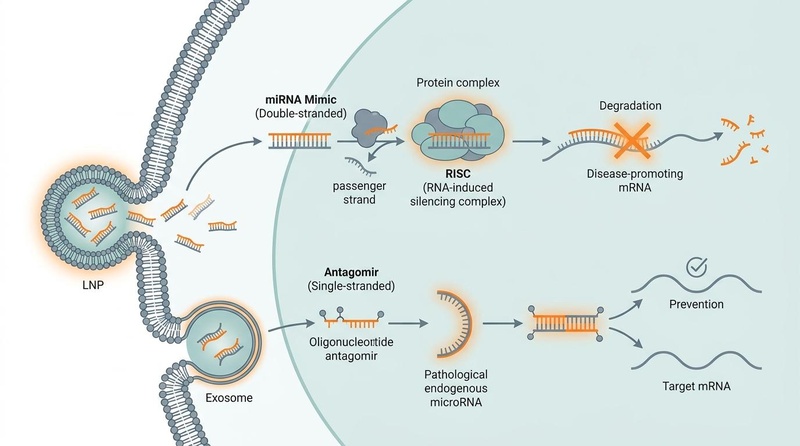
Los microARN funcionan mediante el emparejamiento de bases con los ARNm objetivo. Este mecanismo molecular y su manipulación clínica se detallan en el siguiente esquema:
Fig. 1: Mecanismos clínicos de la terapéutica basada en microARN. A la izquierda, los miméticos de miARN sintéticos de doble cadena se administran a través de nanopartículas lipídicas (LNP), separándose en una cadena pasajera y una cadena guía; la cadena guía se carga en el complejo de silenciamiento inducido por ARN (RISC) para apuntar y degradar el ARNm que promueve la enfermedad. A la derecha, los antagomiRs de oligonucleótidos de cadena sencilla administrados a través de exosomas se unen y secuestran los miARN endógenos maduros patológicos, evitando que silencien los ARNm objetivo.
El mecanismo de acción intracelular está mediado por el complejo de silenciamiento inducido por ARN (RISC). El motor catalítico principal de RISC es la proteína Argonaute (AGO), que recluta la cadena "guía" de miARN de cadena sencilla. Cuando la guía de miARN exhibe una complementariedad perfecta o casi perfecta con el ARNm objetivo (común en plantas y miméticos sintéticos), RISC induce la escisión endonucleolítica directa del transcrito. Más comúnmente en los sistemas de mamíferos, la complementariedad parcial (típicamente en la región 3' UTR) da como resultado la represión traduccional y la posterior deadenilación, acelerando la degradación del ARNm [1:2].
¶ Vías celulares y de longevidad fundamentales
El bucle de senescencia p53/miR-34a/SIRT1
Una de las vías de miARN más estudiadas en el envejecimiento de los mamíferos es el bucle de retroalimentación positiva p53/miR-34a/SIRT1. Bajo estrés celular crónico o daño en el ADN, el supresor de tumores p53 se activa, transcribiendo directamente el microARN prosenescente miR-34a [11:1]. A su vez, miR-34a se une directamente y reprime el ARNm de Sirtuin 1 (SIRT1), una deacetilasa dependiente de NAD+ fundamental que promueve la reparación del ADN, la biogénesis mitocondrial y la supervivencia celular [11:2][16].
A medida que caen los niveles de SIRT1, p53 permanece altamente acetilado y activo, impulsando una mayor transcripción de miR-34a y consolidando la senescencia celular. Romper terapéuticamente este bucle utilizando antagomiRs dirigidos a miR-34a representa una estrategia de primer nivel para rescatar la expresión de SIRT1, restaurar la función mitocondrial y rejuvenecer los nichos de células madre senescentes [11:3][16:1].
Chronic Stress/DNA Damage ──> p53 Activation
│
▼
miR-34a Transcription ──┐
▲ │ (Inhibition)
│ ▼
p53 Acetylation <── SIRT1 Repression
(Highly Active)
Integridad vascular y función endotelial (miR-126)
El envejecimiento vascular es impulsado por una disfunción endotelial progresiva. El endotelio depende en gran medida de miR-126, que mantiene la integridad vascular, suprime la adhesión de leucocitos y promueve la angiogénesis al dirigirse a los inhibidores de la vía de señalización de VEGF (específicamente SPRED1 y PIK3R2) [17]. Durante el envejecimiento cronológico y en enfermedades metabólicas como la diabetes tipo 2, los niveles circulantes y tisulares de miR-126 se pierden drásticamente, lo que conduce a un riesgo cardiovascular acelerado y a una mala recuperación de la isquemia [17:1][18].
Alteraciones epigenéticas y las marcas del envejecimiento
Más allá de los bloqueos traduccionales directos, los miARN interactúan dinámicamente con el panorama epigenético. Los "Epi-miRs" (como la familia miR-29) se dirigen directamente a las ADN metiltransferasas (DNMT3A y DNMT3B), modulando así los patrones globales de metilación del ADN. La pérdida de miR-29 con la edad conduce a la hipermetilación de loci genómicos y a la regulación a la baja de genes protectores de remodelación tisular, lo que ilustra cómo la desregulación de los miARN se propaga a través de múltiples marcas del envejecimiento [6:3][1:3].
¶ Dosis y protocolos
Debido a que las terapias con microARN están en fase de investigación y están sujetas a una rápida eliminación biológica, su aplicación clínica depende en gran medida de modificaciones bioquímicas avanzadas y vehículos de administración dirigida.
¶ Tecnologías de administración y formulaciones
¶ Nanopartículas lipídicas (LNPs)
Comúnmente utilizadas para empaquetar ácidos nucleicos dentro de una envoltura lipídica protectora, las LNPs protegen a los miARN de la degradación enzimática en la circulación y facilitan la entrada celular a través de la endocitosis [4:3]. Sin embargo, las LNPs no dirigidas se acumulan predominantemente en el hígado y el bazo, y pueden desencadenar una activación inmunitaria sistémica grave. La administración sistémica mediada por LNPs de miméticos de miR-34a (MRX34), por ejemplo, se vio limitada por una alta inmunogenicidad y una liberación sistémica tóxica de citoquinas [4:4].
¶ Exosomas (Vesículas Extracelulares)
Vesículas a nanoescala (30–150 nm) de origen natural que transfieren biomoléculas entre células. Los exosomas presentan proteínas de membrana (como CD9, CD63, CD81 e integrinas) que facilitan una administración biocompatible y de baja inmunogenicidad a través de las barreras fisiológicas [9:3][15:1]. Debido a que evitan las respuestas inmunitarias severas desencadenadas por las LNPs sintéticas, los exosomas modificados se consideran el vehículo estándar de oro para la administración de miméticos de miARN antisenescentes y antagomiros [15:2].
¶ Minicélulas Bacterianas Dirigidas (EDVs)
Los EnGeneIC Dream Vectors (EDVs) son minicélulas no viables derivadas de bacterias, recubiertas con anticuerpos biespecíficos específicos de células (por ejemplo, anti-EGFR). Este diseño dirige la carga de la minicélula (como un mimético de miR-16, TargomiRs) directamente a las células objetivo, evitando los tejidos sistémicos y previniendo la toxicidad hepática [14:1].
¶ Oligonucleótidos Antisentido (ASOs) y Modificaciones Químicas
Para prolongar la vida media y mejorar la captación tisular, los antagomiros monocatenarios se modifican con químicas como:
- Locked Nucleic Acids (LNAs): Nucleótidos conformacionalmente restringidos que aumentan drásticamente la estabilidad de hibridación y la temperatura de fusión térmica, lo que permite obtener antagomiros más cortos y altamente estables [7:2][8:3].
- Modificaciones 2'-O-Methyl (2'-O-Me) y 2'-Fluoro (2'-F): Previenen la escisión por nucleasas.
- Enlaces de fosforotioato (PS): Previenen la eliminación rápida por filtración renal.
- Conjugación con colesterol (Chol-conjugated): Facilita el transporte transmembrana directo sin requerir un portador de nanopartículas [6:4].
¶ Paradigmas de Dosificación en Estudios Clínicos
Dado que no existe un régimen de dosificación estandarizado de venta libre para los microARN, los protocolos clínicos y de investigación son específicos para cada indicación:
- Control de Fibrosis Localizada (Remlarsen, mimético de miR-29): Se administra mediante inyecciones intradérmicas locales a una dosis de 3,0 mg a 10,0 mg por centímetro de margen de la herida, repetidas semanalmente durante las fases activas de curación para suprimir la formación de queloides [6:5].
- Control Inflamatorio Sistémico (Obefazimod, regulador al alza de miR-124): Se administra como una dosis oral de 50 mg a 100 mg una vez al día. Esta pequeña molécula regula al alza el miR-124 en las células inmunitarias, suprimiendo las vías inflamatorias de manera sostenida durante 8 a 48 semanas [9:4].
- Administración Sistémica de Antagomiros (Miravirsen, anti-miR-122): Se administra mediante inyecciones subcutáneas en dosis que oscilan entre 3,0 y 7,0 mg/kg, administradas semanalmente durante 5 semanas consecutivas, proporcionando una supresión viral sostenida por hasta 14 semanas debido a la vida media en tejidos profundos de los compuestos modificados con LNA [7:3].
¶ Seguridad
El perfil de seguridad de las terapias con microARN depende en gran medida del vehículo de administración, la precisión de la focalización en los tejidos y el grado de hibridación inespecífica (off-target).
¶ Límite crítico de seguridad: Inmunogenicidad e inflamación
La administración sistémica de miméticos sintéticos de miARN de doble cadena conlleva un alto riesgo de desencadenar el sistema inmunitario innato. En el ensayo de fase I de MRX34 (un mimético de miR-34a encapsulado en LNP), la administración sistémica causó eventos adversos graves y mortales mediados por el sistema inmunitario (incluyendo neumonitis, colitis y síndrome de liberación de citocinas) en múltiples pacientes, lo que resultó en la terminación del ensayo [4:5]. El ARN de doble cadena puede activar los receptores de tipo Toll (TLR3, TLR7, TLR8) y los sensores citosólicos, iniciando una respuesta inflamatoria profunda.
¶ Peligro espaciotemporal: Regeneración incontrolada
Si bien los microARN poseen un notable potencial regenerativo, la expresión continua y no regulada puede tener consecuencias catastróficas. En un estudio histórico de Nature, la administración del miR-199a humano a través de un vector viral (AAV6) estimuló una potente proliferación de cardiomiocitos y la reducción de cicatrices en cerdos tras un infarto de miocardio [5:1]. Sin embargo, debido a que el vector viral no se podía apagar, la expresión se mantuvo activa, lo que llevó a la formación de mioblastos poco diferenciados y altamente arritmogénicos. Esto desencadenó taquiarritmias ventriculares repentinas y letales en el 100 % de los animales tratados a las 7-8 semanas posteriores al tratamiento, lo que ilustra la necesidad absoluta de sistemas de expresión transitorios y estrictamente controlados [5:2].
¶ Hibridación fuera de la diana
Debido a que los miARN requieren solo una complementariedad parcial (principalmente la secuencia semilla de 7 nucleótidos) para unirse y reprimir los transcritos diana, los miméticos sintéticos o los antagomiros en dosis altas pueden unirse inadvertidamente y suprimir ARNm no deseados en tejidos sanos. Este silenciamiento fuera de la diana puede interrumpir vías homeostáticas críticas, lo que subraya la necesidad de una administración altamente localizada o de vectores de direccionamiento específicos de tejido [5:3][1:4].
¶ Indicadores de interrupción terapéutica y monitorización clínica
Los protocolos de investigación que utilizan terapias de ARN sistémicas requieren una monitorización clínica rigurosa para detectar marcadores tempranos de toxicidad. Los indicadores para el cese terapéutico inmediato incluyen:
- Marcadores inflamatorios sistémicos que aumentan rápidamente por encima del valor basal (p. ej., proteína C reactiva > 10 mg/L o elevaciones agudas de IL-6).
- Recuento de plaquetas que cae por debajo de 100.000/μL (indicativo de trombocitopenia inducida por oligonucleótidos).
- Transaminasas hepáticas (ALT/AST) que se elevan a >3 veces el límite superior de la normalidad, lo que indica saturación y toxicidad hepática.
- Síntomas pulmonares inexplicables, disnea o malestar gastrointestinal, que pueden anunciar una neumonitis o colitis mediada por el sistema inmunitario en etapa temprana.
¶ Preguntas frecuentes prácticas
¿Cuál es la diferencia entre un siRNA y un microRNA?
Aunque ambos son ARN pequeños no codificantes, los ARN de interferencia pequeños (siRNAs) suelen ser de origen exógeno, diseñados con una complementariedad de secuencia del 100% para dirigirse a un único transcrito de mRNA específico y silenciarlo. Por el contrario, los microRNAs son reguladores endógenos diseñados con complementariedad parcial, lo que les permite dirigirse a redes enteras de docenas o cientos de mRNAs simultáneamente y coordinarlas [1:5].
¿Se pueden utilizar los microRNAs circulantes para medir mi edad biológica?
Sí. Los estudios clínicos y de cohortes han validado paneles específicos de microRNAs circulantes como biomarcadores de envejecimiento altamente sensibles. Los modelos de edad biológica entrenados con perfiles de expresión de microRNAs (como mirAge, mirMort y mirFI) se correlacionan sólidamente con el funcionamiento físico, la fragilidad y la mortalidad por todas las causas a 10 años, superando a la edad cronológica en la predicción de riesgos [3:3].
¿Por qué no puedo comprar suplementos de microRNA en línea?
Los microRNAs sintéticos son altamente inestables y son degradados rápidamente por las nucleasas de ARN en la sangre en cuestión de minutos si no están protegidos por modificaciones químicas o vectores de administración complejos [4:6]. Además, la administración sistémica conlleva altos riesgos de inmunogenicidad y efectos graves fuera del objetivo (off-target). Por lo tanto, las terapias con microRNA siguen restringidas a ensayos clínicos de investigación bajo un estricto seguimiento [4:7][5:4].
¿Cómo mejoran los exosomas la administración de terapias con microRNA?
Los exosomas son vesículas unidas a membranas y biocompatibles producidas de forma natural por las células humanas. Debido a que expresan proteínas de superficie nativas, pueden eludir el reconocimiento inmunológico, evitar la rápida eliminación por parte del hígado y administrar los miRNAs directamente en los tejidos diana. Esto los convierte en portadores significativamente más seguros y eficaces que las nanopartículas lipídicas (LNPs) sintéticas [9:5][15:3].
¿Existe alguna forma de modular mis microRNAs de forma natural?
En estudios observacionales se ha demostrado que ciertas intervenciones en el estilo de vida, como el ejercicio físico regular y la restricción calórica, restauran parcialmente los perfiles de expresión de microRNAs juveniles, como la regulación al alza del miR-126 protector y la regulación a la baja de los inflamma-miRs proinflamatorios, aunque estos cambios son modestos en comparación con las intervenciones terapéuticas directas [1:6][18:1].
¶ Referencias
Kinser HE, Pincus Z. MicroRNAs as modulators of longevity and the aging process. Hum Genet. 2020;139(1):17-31. https://pmc.ncbi.nlm.nih.gov/articles/PMC6954352/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Grillari J, et al. Profiling microRNA expression during senescence and aging: mining for a diagnostic tool of senescent-cell burden. bioRxiv. 2024;2024.04.10.588794. https://www.biorxiv.org/content/10.1101/2024.04.10.588794v2 ↩︎ ↩︎ ↩︎
Rotterdam Study Consortium. Plasma microRNA signatures of aging and their links to health outcomes and mortality: findings from a population-based cohort study. Genome Med. 2025;17(1):15. https://pmc.ncbi.nlm.nih.gov/articles/PMC12188677/ ↩︎ ↩︎ ↩︎ ↩︎
Beg MS, Brenner AJ, Sachdev J, et al. Phase I Study of MRX34, a Liposome-Formulated mR-34a Mimic, in Patients With Advanced Solid Tumors. J Clin Oncol. 2019;37(14):1128-1136. https://ascopubs.org/doi/full/10.1200/JCO.18.01254 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Gabisonia K, Prosdocimo G, Diez-Ciudad I, et al. MicroRNA therapy stimulates uncontrolled cardiac repair after myocardial infarction in pigs. Nature. 2019;575(7782):356-362. https://www.nature.com/articles/s41586-019-1191-6 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Gallant-Behm CL, Zhang Y, Lee SJ, et al. A MicroRNA-29 Mimic (Remlarsen) Represses Extracellular Matrix Expression and Fibroplasia in the Skin. J Invest Dermatol. 2019;139(3):614-623. https://www.sciencedirect.com/science/article/pii/S0022202X18328306 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Janssen HL, Reesink HW, Lawitz EJ, et al. Treatment of HCV Infection by Targeting MicroRNA. N Engl J Med. 2013;368(18):1685-1694. https://www.nejm.org/doi/full/10.1056/NEJMoa1209026 ↩︎ ↩︎ ↩︎ ↩︎
Foss F, et al. Phase 1 Trial of Cobomarsen (MRG-106), an Inhibitor of microRNA-155, in Patients with Cutaneous T-Cell Lymphoma. Blood. 2018;132(Suppl 1):2902. https://ashpublications.org/blood/article/132/supplement 1/2902/263360/phase-1-trial-of-cobomarsen-mrg-106-an-inhibitor ↩︎ ↩︎ ↩︎ ↩︎
Vermeire S, et al. ABX464 (Obefazimod) Upregulates miR-124 to Reduce Proinflammatory Markers in Inflammatory Bowel Diseases. Clin Transl Gastroenterol. 2023;14(4):e00572. https://pmc.ncbi.nlm.nih.gov/articles/PMC10132720/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Noureddine M, et al. Circulating microRNA profile of long-lived Okinawans identifies novel potential targets for optimizing lifespan and health span. Aging Cell. 2024;23(8):e14191. https://onlinelibrary.wiley.com/doi/10.1111/acel.14191 ↩︎ ↩︎
Yamakuchi M, Lowenstein CJ. MiR-34, SIRT1 and p53: the feedback loop. Cell Cycle. 2009;8(5):712-713. https://pubmed.ncbi.nlm.nih.gov/19221490/ ↩︎ ↩︎ ↩︎ ↩︎
Abplanalp WT, Fischer A, John D, et al. Efficiency and Target Derepression of Anti-miR-92a: Results of a First in Human Study. Nucleic Acid Therapeutics. 2020;30(6):335-342. https://journals.sagepub.com/doi/abs/10.1089/nat.2020.0871 ↩︎
Gallant-Behm CL, Piper J, Dickinson BA, et al. A synthetic microRNA-92a inhibitor (MRG-110) accelerates angiogenesis and wound healing in diabetic and nondiabetic wounds. Wound Repair Regen. 2018;26(4):311-323. https://pubmed.ncbi.nlm.nih.gov/30118158/ ↩︎
van Zandwijk N, Pavlakis N, Kao SC, et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: a first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017;18(11):1456-1466. https://www.thelancet.com/journals/lanonc/article/PIIS1470-2045(17)30621-6/abstract ↩︎ ↩︎
Ipson BR, et al. Identifying exosome-derived microRNAs as candidate biomarkers of frailty. Sci Rep. 2018;8(1):15339. https://pmc.ncbi.nlm.nih.gov/articles/PMC6384524/ ↩︎ ↩︎ ↩︎ ↩︎
Zhao T, Li Z, Pang S, et al. P53/miR-34a/SIRT1 positive feedback loop regulates the termination of liver regeneration. Aging (Albany NY). 2021;13(4):5062-5079. https://www.aging-us.com/article/203920/text ↩︎ ↩︎
Zampetaki A, Kiechl S, Drozdov I, et al. Plasma MicroRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ Res. 2010;107(6):810-817. https://doi.org/10.1161/CIRCRESAHA.110.226357 ↩︎ ↩︎
de Lucia C, et al. MicroRNA-based therapeutics in cardiovascular disease. Eur J Intern Med. 2017;43:10-15. https://doi.org/10.1016/j.ejim.2017.05.011 ↩︎ ↩︎