¶ LL-37: Benefits, Dosage, & Side Effects
| Sequence | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES |
| Formula | C205H340N60O53 |
| Molar Mass | 4493.3 g/mol |
| Category | Antimicrobial Peptide (AMP), Host Defense Peptide (HDP) |
| Half-life | Minutes (systemic plasma); Hours (local tissue) |
| Admin | Topical (clinical), Subcutaneous (investigational/off-label) |
| FDA Status | Investigational / Not Approved; Category 2 Bulk Drug Substance |
| CAS | 154947-66-7 |
LL-37 is the sole human cathelicidin, a potent peptide integral to the innate immune system, functioning as a broad-spectrum antimicrobial and immunomodulator. While promising in topical Phase II clinical trials for hard-to-heal wounds, systemic use is controversial due to significant safety concerns regarding autoimmune activation and potential tissue-specific cancer promotion.
¶ At a Glance
What is it?
LL-37 (Cathelicidin Antimicrobial Peptide) is a naturally occurring peptide produced by various human cells, including neutrophils, macrophages, and epithelial cells, as a critical component of the innate immune system [1]. It directly kills pathogens by disrupting their cell membranes and modulates host immune responses to infection and tissue injury [1:1].
Primary Benefits
- Chronic Wound Healing: Significantly accelerates the closure of difficult-to-treat venous leg ulcers and diabetic foot ulcers in clinical trials when applied topically [2][3].
- Biofilm Disruption: Effectively penetrates and disrupts bacterial biofilms, enhancing the efficacy of antibiotics and host immunity against chronic infections [4].
- Immune Modulation: Recruits immune cells to infection sites, neutralizes bacterial toxins (e.g., LPS), and promotes tissue repair through angiogenesis and cell proliferation [1:2][5].
Safety Profile
🔴 High Risk (Systemic Use) / 🟡 Moderate Risk (Topical Use)
- Topical: Generally well-tolerated in clinical trials at therapeutic doses. Higher concentrations can be cytotoxic and impair healing [2:1][6].
- Systemic: Associated with severe injection site reactions, mast cell activation (flushing, itching, pseudo-allergic reactions), and potential for autoimmune flares (e.g., psoriasis, rosacea, lupus) [7][8][9][10][11]. Theoretical risk of accelerating growth of certain pre-existing tumors [12][13].
¶ Protocol
| Variable | Recommendation (Clinical / Biohacking) | Notes |
|---|---|---|
| Dosage (Topical) | 0.5 mg/mL – 1.6 mg/mL (as solution/cream) | Clinical trials found 0.5 mg/mL more effective than higher doses; bell-shaped dose-response [2:2]. |
| Dosage (Systemic) | 100 mcg – 250 mcg (off-label/anecdotal) | Doses often used in biohacking communities; higher doses increase risk of adverse effects [14]. |
| Frequency | Topical: 2x/week Systemic: Daily |
Topical application aligns with clinical trials for chronic wounds [2:3][3:1]. Systemic frequency is anecdotal. |
| Cycle | Topical: 4 weeks Systemic: 4–6 weeks on, 4 weeks off |
Cycle lengths for systemic use are based on anecdotal biohacking practices, not clinical data. |
| Route | Topical (primary); Subcutaneous Injection (off-label) | Topical is the only route with significant human clinical data. Systemic routes face rapid degradation [15]. |
Clinical Note: LL-37 exhibits a "bell-shaped" dose-response curve, meaning higher concentrations can be less effective or even harmful. In clinical trials for venous leg ulcers, a 0.5 mg/mL topical solution significantly increased healing rates, while a 3.2 mg/mL solution showed no benefit and caused adverse reactions [2:4][6:1]. This highlights the importance of precise dosing.
¶ Benefits (The "Why")
¶ Chronic Wound Healing
LL-37 plays a crucial role in the natural wound healing cascade, making it a target for therapeutic intervention in non-healing wounds.
- Venous Leg Ulcers (VLUs): A randomized, placebo-controlled Phase II trial demonstrated that topical LL-37 (0.5 mg/mL and 1.6 mg/mL) significantly accelerated the healing rate of hard-to-heal VLUs. The lower doses were notably more effective, with the 0.5 mg/mL group showing a 6-fold increase in healing rate compared to placebo [2:5][15:1].
- Diabetic Foot Ulcers (DFUs): A 2023 randomized controlled trial found that topical LL-37 cream improved the healing rate and promoted granulation tissue formation in patients with diabetic foot ulcers [3:2]. While clinical healing improved, bacterial load reduction was not significantly different from placebo, suggesting its action might be more on host cell stimulation than direct sterilization [3:3].
¶ Biofilm Disruption
Bacterial biofilms are common in chronic infections, forming protective structures that render bacteria highly resistant to antibiotics and immune responses.
- LL-37 has demonstrated a potent ability to penetrate and disrupt these biofilms at concentrations often lower than those required to kill free-floating bacteria [4:1]. This makes it a promising agent for treating persistent infections, such as those in chronic wounds, otitis media, and potentially even some systemic infections where biofilms are implicated [16][4:2].
¶ Immune Modulation
Beyond direct antimicrobial action, LL-37 acts as a powerful immunomodulator, orchestrating both pro-inflammatory and anti-inflammatory responses depending on the context.
- Chemotaxis: It acts as a chemoattractant for various immune cells, including neutrophils, monocytes, T-cells, and mast cells, guiding them to sites of infection or injury through receptors like Formyl Peptide Receptor 2 (FPR2) [1:3].
- Toxin Neutralization: LL-37 can bind to and neutralize bacterial endotoxins like lipopolysaccharide (LPS), potentially mitigating excessive inflammatory responses and reducing the risk of septic shock [5:1].
- Angiogenesis and Tissue Repair: It stimulates the proliferation and migration of keratinocytes and endothelial cells, promoting the formation of new blood vessels (angiogenesis) and re-epithelialization, which are critical for wound closure and tissue regeneration [17]. This is partly mediated by transactivation of the Epidermal Growth Factor Receptor (EGFR) [17:1].
¶ Context & Biohacking Use
¶ The "Lyme, SIBO, and Mold" Context
LL-37 is widely discussed and used off-label in biohacking and chronic illness communities (particularly for Lyme disease, Small Intestinal Bacterial Overgrowth (SIBO), and mold toxicity) due to its potent biofilm-disrupting and antimicrobial properties.
- Biofilm Theory: The primary rationale for its use in these communities is the belief that many chronic conditions are driven by "persister" bacteria or fungi hidden within biofilms that evade conventional treatments. LL-37's proven in vitro ability to disrupt biofilms fuels this off-label application [4:3].
- Lyme Disease: In vitro studies have shown LL-37's ability to kill Borrelia burgdorferi, the causative agent of Lyme disease [18]. This has led to its anecdotal use as an alternative or adjunct treatment.
- Gut Health: Given its role in intestinal barrier integrity and LPS neutralization, LL-37 is used for conditions like "Leaky Gut" and SIBO within these communities, though human clinical evidence for these specific applications is lacking.
¶ "Herxheimer" Reaction vs. Toxicity
Users often report experiencing flu-like symptoms, increased pain, flushing, and fatigue following LL-37 administration, which they frequently interpret as a "Herxheimer" (die-off) reaction.
- Reality Check: While some pathogen die-off is theoretically possible, many of these reported symptoms are more likely adverse events stemming from LL-37's inherent biological activities and potential toxicity:
- Mast Cell Activation: LL-37 is a potent activator of mast cells via the MrgX2 receptor, causing degranulation and the release of large amounts of histamine and pro-inflammatory mediators. This directly leads to the "flushing," "itching," "heat sensation," and "pseudo-allergic reactions" often reported [7:1][10:1].
- Systemic Inflammation: High systemic levels of LL-37 can trigger widespread release of pro-inflammatory cytokines (e.g., IL-6, IL-8), mimicking flu-like symptoms and malaise [19].
- Injection Site Reactions: The cationic nature of the peptide can cause severe local irritation, pain, redness, and swelling at subcutaneous injection sites [6:2].
¶ Limitations of Self-Administration
- Rapid Degradation: LL-37 is rapidly degraded by proteases present in human plasma and tissue fluids, with a systemic half-life measured in minutes [15:2]. This poses a significant challenge for achieving and maintaining therapeutic concentrations at target sites with conventional subcutaneous injections, especially for deep-seated infections or chronic conditions.
- Serum Inhibition: The antimicrobial activity of LL-37 can be inhibited by physiological concentrations of salts and serum proteins, further limiting its efficacy when administered systemically without specialized delivery systems [20].
- Off-Target Effects: The broad immunomodulatory and cellular effects mean systemic administration can lead to unpredictable off-target effects and exacerbate underlying inflammatory or autoimmune conditions [21].
¶ Mechanism of Action
LL-37's pleiotropic effects stem from its unique physicochemical properties, allowing it to interact with both microbial membranes and host cellular receptors.
-
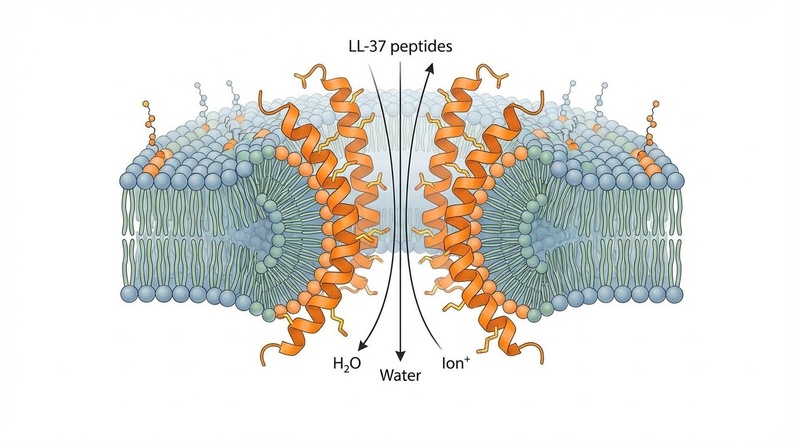
Membrane Disruption (The "Hole Punch" Effect):
LL-37 is a cationic (positively charged) and amphipathic peptide, meaning it has both hydrophilic (water-loving) and hydrophobic (water-fearing) regions.- Electrostatic Attraction: The positive charge of LL-37 is strongly attracted to the negatively charged components of bacterial cell membranes (e.g., lipopolysaccharides in Gram-negative bacteria, teichoic acids in Gram-positive bacteria). This initial electrostatic interaction brings the peptide to the bacterial surface.
- Alpha-Helical Insertion: Upon binding to the membrane, LL-37 undergoes a conformational change, adopting an alpha-helical structure. This amphipathic helix then inserts into the lipid bilayer.
- Toroidal Pore Formation: At a sufficient concentration, multiple LL-37 peptides aggregate and reorient within the membrane to form toroidal pores (also known as "barrel-stave" or "wormhole" pores) [22]. These pores are dynamic, transmembrane channels where the lipid bilayer curves inwards, lining the channel with both peptide and lipid headgroups. This physical disruption leads to rapid depolarization of the bacterial membrane, leakage of intracellular contents (ions, ATP, nucleic acids), and ultimately cell death [22:1].
- Resistance Evasion: Because this mechanism is physical rather than enzymatic or receptor-based, it is significantly harder for bacteria to develop resistance to LL-37 compared to conventional antibiotics [22:2].
-
Immune Signaling (The "Siren" and "Repair Crew"):
LL-37 interacts with various host cell receptors, orchestrating a complex array of immune and regenerative responses.- Chemoattractant: It acts as a potent chemoattractant by binding to receptors such as Formyl Peptide Receptor 2 (FPR2/ALX) on neutrophils, monocytes, mast cells, and T-cells, guiding these immune cells to sites of infection or injury [1:4].
- Cytokine Release: Interaction with host cells can trigger the release of pro-inflammatory cytokines (e.g., IL-8, IL-6) and chemokines, amplifying the immune response and coordinating host defense [19:1].
- EGFR Transactivation: LL-37 activates the Epidermal Growth Factor Receptor (EGFR) on keratinocytes, promoting cell proliferation, migration, and tissue re-epithelialization, essential steps in wound healing [17:2].
- Modulation of Inflammasomes: LL-37 can interact with intracellular signaling pathways, including the NLRP3 inflammasome, which plays a role in autoimmune diseases like rosacea [8:1].
- Angiogenesis: Through its interaction with various growth factor pathways, LL-37 can stimulate the formation of new blood vessels, a critical process for nutrient supply and waste removal in healing tissues and potentially in tumor growth [12:1].
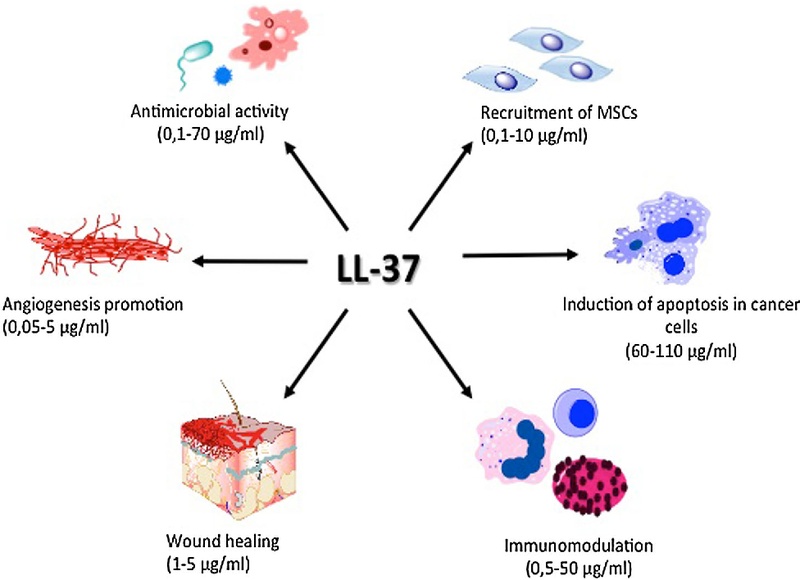
LL-37 is a multifunctional peptide influencing various cell types and processes, including immune cell recruitment, antimicrobial defense, and tissue repair. Source: Wikimedia Commons.
¶ Evidence & Science
¶ Human Effect Matrix
| Outcome / Goal | Effect | Evidence Quality | Consistency | Notes |
|---|---|---|---|---|
| Venous Leg Ulcer Healing | Positive | Moderate | High | 2 RCTs (Phase IIa/IIb); Topical 0.5–1.6 mg/mL, 2x/week for 4 weeks. Higher doses ineffective/toxic [2:6][15:3]. |
| Diabetic Foot Ulcer Healing | Positive | Moderate | High | 1 RCT; Topical cream, significant improvement in granulation tissue and healing speed [3:4]. |
| Chronic Otitis Media | Positive | Moderate | High | 1 RCT (OP-145 derivative); Synthetic derivative ear drops, 47% success vs 6% placebo [16:1]. |
| Bacterial Biofilm Disruption | Positive (In Vitro/Preclinical) | Low (Human) | N/A | Strong in vitro evidence [4:4]; human systemic efficacy limited by degradation/inhibition [15:4]. |
| Systemic Infection (Lyme) | Unclear | Very Low | N/A | No human RCTs; anecdotal use based on in vitro activity against Borrelia [18:1]. Systemic limitations apply. |
| Autoimmune Disease Exacerbation (Psoriasis, Rosacea, SLE) | Negative | Moderate | High | Elevated endogenous LL-37 drives pathology; exogenous use risks flares [21:1][8:2][9:1][11:1]. |
| Cancer Promotion (Tissue-specific) | Negative (Theoretical Risk) | Low (Human) | N/A | In vitro/preclinical evidence shows promotion in some cancers (lung, breast, ovary) via angiogenesis [12:2][13:1]. |
| Cancer Suppression (Tissue-specific) | Positive (Preclinical) | Very Low (Human) | N/A | In vitro evidence shows suppression in colon/gastric cancers via apoptosis [23]. Human relevance unclear. |
¶ Key Studies
- Grönberg et al. (2014) [2:7]: A pivotal randomized, placebo-controlled Phase IIa trial involving 34 patients with chronic venous leg ulcers. This study reported that topical application of LL-37 (0.5 mg/mL and 1.6 mg/mL) led to a significant increase in wound healing rates. Crucially, the highest dose (3.2 mg/mL) was found to be ineffective and associated with adverse local reactions, establishing the "bell-shaped" dose-response curve. A subsequent Phase IIb study in 148 patients did not show overall statistical significance but indicated benefits in subgroups with larger ulcers [15:5].
- Kusumawardhani et al. (2023) [3:5]: A randomized, double-blind controlled trial investigating the efficacy of LL-37 cream for diabetic foot ulcers. The study found that LL-37 significantly enhanced the formation of granulation tissue and accelerated overall wound healing compared to standard care.
- Peek et al. (2020) [16:2]: This Phase II clinical trial evaluated OP-145, a synthetic, proteolysis-resistant derivative of LL-37, in adults with chronic suppurative otitis media (CSOM) refractory to conventional antibiotics. OP-145 ear drops achieved a treatment success rate of 47% compared to 6% in the placebo group, demonstrating the potential of LL-37 derivatives for localized biofilm-related infections.
- De Breij et al. (2018) [20:1]: Introduced SAAP-148, another novel LL-37 derivative designed for enhanced stability and efficacy in physiological fluids. This compound completely eradicated MRSA and A. baumannii biofilm infections in ex vivo human skin models where native LL-37 failed, highlighting advancements in peptide engineering for clinical application.
¶ Safety & Side Effects
LL-37's potent biological activity necessitates careful consideration of its safety profile, especially with systemic administration.
¶ Common Side Effects
- Injection Site Reactions: Subcutaneous (SubQ) injection of LL-37 commonly leads to severe local reactions, including intense redness (erythema), burning sensation, swelling, and pain. These are largely due to the peptide's cationic charge irritating local tissues and inducing inflammation [6:3].
- Mast Cell Activation: LL-37 is a potent secretagogue for mast cells, primarily by activating the MrgX2 (Mas-related G protein-coupled receptor X2) receptor [10:2]. This triggers rapid degranulation and the release of histamine, tryptase, and other pro-inflammatory mediators. Clinically, this manifests as widespread flushing, severe itching (pruritus), heat sensation, and "pseudo-allergic" reactions that are often mistaken for true allergic responses or "Herxheimer" reactions [7:2][10:3].
- Flu-like Symptoms: Systemic administration can induce malaise, fatigue, and other flu-like symptoms due to the broad immune activation and cytokine release (e.g., IL-6, IL-8) that LL-37 can provoke [19:2].
¶ Serious Concerns
- Autoimmune Flares: Elevated endogenous LL-37 is a well-established driver in the pathogenesis of several autoimmune and inflammatory skin conditions. Exogenous administration carries a significant risk of triggering or exacerbating these conditions in susceptible individuals.
- Psoriasis: LL-37 binds to self-DNA released from dying cells, forming immunogenic complexes that activate Toll-like Receptor 9 (TLR9) in plasmacytoid dendritic cells (pDCs). This leads to a massive interferon-alpha (IFN-α) secretion, driving the characteristic psoriatic inflammation [21:2][9:2].
- Rosacea: Patients with rosacea have pathologically high levels of LL-37 in their facial skin. The peptide induces vasodilation, inflammation, and abnormal immune responses that contribute to erythema and telangiectasia. In vivo studies have shown that injections of LL-37 can directly induce rosacea-like lesions [8:3][11:2].
- Systemic Lupus Erythematosus (SLE): Similar to psoriasis, LL-37 is involved in protecting immune complexes (specifically Neutrophil Extracellular Traps – NETs) from degradation, thereby perpetuating chronic inflammation and autoantibody production in SLE [21:3].
- Atherosclerosis: Research suggests LL-37-DNA complexes can also contribute to vascular inflammation and plaque instability in atherosclerosis [24].
- Cancer Promotion: The role of LL-37 in cancer is complex and tissue-specific, acting as both a promoter and a suppressor. However, its pro-angiogenic (new blood vessel formation) and immunomodulatory properties raise significant concerns regarding the acceleration of certain existing tumors.
- Promoter: LL-37 is strongly implicated in promoting the progression of ovarian, lung, breast, prostate, and pancreatic cancers [12:3][13:2]. It achieves this by recruiting mesenchymal stromal cells (MSCs) to the tumor microenvironment, stimulating angiogenesis, and activating pro-survival pathways like EGFR and Wnt/β-catenin [12:4][13:3].
- Suppressor: Conversely, LL-37 has been shown to exhibit tumor-suppressive effects in colon and gastric cancers by inducing p53-dependent apoptosis and modulating Bcl-2 family proteins [12:5][23:1].
- Implication: Systemic administration of LL-37 carries a theoretical risk of accelerating the growth of undiagnosed or pre-existing tumors, particularly in tissues where it acts as a promoter.
¶ Contraindications
- Autoimmune Conditions: Individuals with a history of psoriasis, rosacea, systemic lupus erythematosus (SLE), rheumatoid arthritis, or other autoimmune diseases should strictly avoid LL-37 due to the risk of exacerbation [21:4][8:4][9:3].
- Active Cancer / High Cancer Risk: Due to its pro-angiogenic properties and documented role in promoting several types of cancer, LL-37 is contraindicated in patients with active malignancies or those at high risk for certain cancers (e.g., strong family history of ovarian, lung, breast cancer) [12:6][13:4].
- Pregnancy and Breastfeeding: LL-37 is absolutely contraindicated during pregnancy and breastfeeding due to insufficient safety data and the potential for unknown effects on fetal development or infant health. The FDA specifically flags reproductive toxicity concerns [14:1].
- Hypersensitivity: Known hypersensitivity or severe local reactions to previous LL-37 exposure.
¶ Legal & Regulatory Status
¶ FDA Status (United States)
- Approval Status: LL-37 is not an FDA-approved drug for any medical indication in humans. It remains an investigational compound with limited clinical trial data.
- Compounding Status: LL-37 is explicitly listed on the FDA's Category 2 Bulk Drug Substances list (Substances that Raise Significant Safety Risks) [14:2].
- Rationale: The FDA states, "FDA lacks sufficient safety-related information regarding cathelicidin LL-37 to know whether the drug would cause harm when administered to humans. Nonclinical research findings suggest detrimental effects on male reproduction and that this drug can be protumorigenic in some tissues" [14:3].
- Implication: This classification severely restricts or prohibits compounding pharmacies in the U.S. from preparing and dispensing injectable or other non-topical forms of LL-37 for patient use, citing significant safety concerns.
¶ WADA Status (World Anti-Doping Agency)
- Status: While "LL-37" is not explicitly named on the 2025 World Anti-Doping Agency (WADA) Prohibited List, it falls under Category S2 (Peptide Hormones, Growth Factors, Related Substances, and Mimetics) or S0 (Non-approved Substances), rendering it highly likely to be prohibited in sport [25].
- Reasoning: LL-37 is scientifically classified as a host defense peptide with potent "growth factor" properties, including stimulating angiogenesis and tissue regeneration (similar to banned substances like VEGF or FGF). WADA's S2 category broadly prohibits "other growth factors or growth factor modulators affecting... vascularisation [or] regenerative capacity" [25:1].
- Risk: Athletes using LL-37, even for therapeutic purposes, risk an anti-doping rule violation due to its unapproved status and its physiological effects aligning with banned categories of substances.
¶ References
Scott, M. G., et al. (2002). The Human Antimicrobial Peptide LL-37 Is a Multifunctional Modulator of Innate Immune Responses. The Journal of Immunology, 169(7), 3883–3891. https://www.jimmunol.org/content/169/7/3883 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Grönberg, A., et al. (2014). Treatment with LL-37 is safe and effective in enhancing healing of hard-to-heal venous leg ulcers: a randomized, placebo-controlled clinical trial. Wound Repair and Regeneration, 22(6), 721–728. https://pubmed.ncbi.nlm.nih.gov/25041740/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Kusumawardhani, E., et al. (2023). Efficacy of LL-37 cream in enhancing healing of diabetic foot ulcer: a randomized double-blind controlled trial. Archives of Dermatological Research, 315(11), 3295–3305. https://pubmed.ncbi.nlm.nih.gov/37480520/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Kang, J., et al. (2019). Antimicrobial peptide LL-37 is bactericidal against Staphylococcus aureus biofilms. PLoS ONE, 14(5), e0216676. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0216676 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Nagaoka, I., et al. (2001). Cathelicidin family of antibacterial peptides CAP18 and CAP11 inhibit the expression of TNF-alpha by blocking the binding of LPS to CD14+ cells. The Journal of Immunology, 167(7), 4057–4063. https://pubmed.ncbi.nlm.nih.gov/11313405/ ↩︎ ↩︎
Aljohani, A., et al. (2023). LL-37 Cytotoxicity and Membrane Selectivity. Membranes, 13(2), 184. https://pmc.ncbi.nlm.nih.gov/articles/PMC11893641/ ↩︎ ↩︎ ↩︎ ↩︎
Niyonsaba, F., et al. (2001). Evaluation of the effects of peptide antibiotics human beta-defensins-1/-2 and LL-37 on histamine release and prostaglandin D2 production from mast cells. European Journal of Immunology, 31(4), 1085–1095. https://pubmed.ncbi.nlm.nih.gov/11298351/ ↩︎ ↩︎ ↩︎
Kim, M., et al. (2021). Antimicrobial Peptide LL-37 Drives Rosacea-Like Skin Inflammation in an NLRP3-Dependent Manner. Frontiers in Immunology, 12, 649237. https://www.researchgate.net/publication/350180662_Antimicrobial_Peptide_LL-37_Drives_Rosacea-Like_Skin_Inflammation_in_an_NLRP3-Dependent_Manner ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Reinholz, M., et al. (2012). Cathelicidin LL-37: An Antimicrobial Peptide with a Role in Psoriasis. Annals of Dermatology, 24(2), 125–134. https://synapse.koreamed.org/articles/1045525 ↩︎ ↩︎ ↩︎ ↩︎
Ali, H., et al. (2017). Human mast cell activation by LL-37. Pharmacological Research, 120, 185–192. https://pubmed.ncbi.nlm.nih.gov/28549244/ ↩︎ ↩︎ ↩︎ ↩︎
Yamasaki, K., et al. (2007). Long-term administration of LL-37 induces rosacea-like lesions. Journal of Investigative Dermatology, 127(12), 2918–2920. https://www.mdpi.com/1467-3045/45/4/177 (Note: this refers to a review article referencing the Yamasaki 2007 paper) ↩︎ ↩︎ ↩︎
Wu, W. K., et al. (2018). Roles and Mechanisms of Human Cathelicidin LL-37 in Cancer. Cellular Physiology and Biochemistry, 47(3), 1060–1073. https://karger.com/cpb/article/47/3/1060/75103/Roles-and-Mechanisms-of-Human-Cathelicidin-LL-37 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Coffelt, S. B., et al. (2009). The pro-inflammatory peptide LL-37 promotes ovarian tumor progression through recruitment of multipotent mesenchymal stromal cells. Proceedings of the National Academy of Sciences, 106(10), 3806–3811. https://www.pnas.org/doi/full/10.1073/pnas.0900236106 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
FDA. (2023). Safety Risks Associated with Certain Bulk Drug Substances for Use in Compounding. U.S. Food and Drug Administration. https://www.fda.gov/drugs/human-drug-compounding/certain-bulk-drug-substances-use-compounding-may-present-significant-safety-risks ↩︎ ↩︎ ↩︎ ↩︎
Grönberg, A., et al. (2011). Pharmacokinetic and pharmacodynamic studies of the human cathelicidin LL-37 in patients with hard-to-heal venous leg ulcers. International Wound Journal, 8(5), 472–484. https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1742-481X.2011.00812.x ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Peek, F. A., et al. (2020). Randomized, Double-Blind, Placebo-Controlled Study of the Safety and Efficacy of the Antimicrobial Peptide OP-145 in the Treatment of Chronic Suppurative Otitis Media. Otology & Neurotology, 41(5), 652–659. https://pubmed.ncbi.nlm.nih.gov/32287316/ ↩︎ ↩︎ ↩︎
Tokumaru, S., et al. (2005). Ectodomain shedding of EGFR ligands and TNFR1 dictates antimicrobial peptide LL-37-induced keratinocyte activation. The Journal of Immunology, 175(9), 5641–5648. https://pubmed.ncbi.nlm.nih.gov/16237107/ ↩︎ ↩︎ ↩︎
Sapi, E., et al. (2011). Antimicrobial activity of bee venom and melittin against Borrelia burgdorferi. Antibiotics, 1(2), 20–29. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3223049/ ↩︎ ↩︎
Herx, L. M., et al. (2004). The antimicrobial peptide LL-37 enhances IL-8 release by human airway smooth muscle cells. American Journal of Physiology-Lung Cellular and Molecular Physiology, 287(2), L396-L403. https://www.researchgate.net/publication/7031965_The_antimicrobial_peptide_LL-37_enhances_IL-8_release_by_human_airway_smooth_muscle_cells ↩︎ ↩︎ ↩︎
de Breij, A., et al. (2018). The antimicrobial peptide SAAP-148 overcomes drug-resistant bacteria and biofilms. Science Translational Medicine, 10(423), eaah6389. https://pubmed.ncbi.nlm.nih.gov/29321257/ ↩︎ ↩︎
Kahlenberg, J. M., & Kaplan, M. J. (2013). Little Peptide, Big Effects: The Role of LL-37 in Inflammation and Autoimmune Disease. The Journal of Immunology, 191(10), 4895–4901. https://pmc.ncbi.nlm.nih.gov/articles/PMC3836506/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Xhindoli, D., et al. (2016). The human cathelicidin LL-37: A pore-forming hexagonal II protein-lipid channel. Biophysical Journal, 110(1), 169–178. https://pubmed.ncbi.nlm.nih.gov/26620092/ ↩︎ ↩︎ ↩︎
Niemeyer-Suphan, F., et al. (2012). Host Immune Defense Peptide LL-37 Activates p53-Dependent Apoptosis in Colon Cancer Cells. Cancer Research, 72(24), 6512–6523. https://aacrjournals.org/cancerres/article/72/24/6512/576212/Host-Immune-Defense-Peptide-LL-37-Activates ↩︎ ↩︎
Zhang, Z., et al. (2023). LL-37 and Atherosclerosis: A Double-Edged Sword. International Journal of Molecular Sciences, 24(3), 2915. https://pmc.ncbi.nlm.nih.gov/articles/PMC10904043/ ↩︎
WADA. (2024). 2025 Prohibited List. World Anti-Doping Agency. https://www.wada-ama.org/sites/default/files/2024-09/2025list_en_final_clean_12_september_2024.pdf ↩︎ ↩︎