¶ Exercise
每个人都会告诉你运动有益健康。但这是为什么呢?让我们一探究竟。
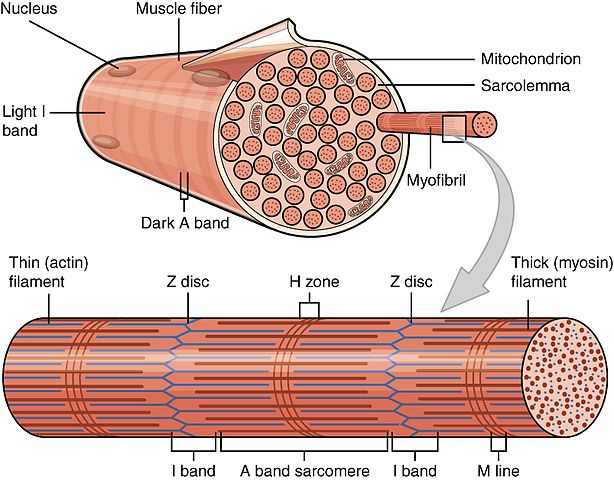
展示肌纤维组织的科学插图,突出了本文讨论的不同纤维类型。
第一个原因是运动能刺激肌肉的生成和维持,肌肉会随着我们年龄的增长而流失,而它对于保持自主能力和生活质量至关重要。
第二个原因是骨骼肌是一个内分泌器官,它分泌调节全身代谢的肌肉因子(myokines)。衰老会优先侵蚀快肌纤维和神经肌肉功能;而训练可以抵消这些变化 [1][2][3][4][5][6]。
¶ Why exercise is healthy (mechanisms, table‑first)

一张科学插图,展示了运动如何触发骨骼肌释放肌肉因子,说明了身体对身体活动的复杂代谢反应。
| 机制 | 主要作用 | 关键结果 | 证据 |
|---|---|---|---|
| 保持肌肉质量和力量 | 减缓肌少症(与年龄相关的肌肉流失) | 维持活动能力,降低残疾和死亡风险 | 观察性队列研究;伴有肥大/力量增加的随机对照试验(RCTs) [4:1][5:1][6:1] |
| 内分泌信号传导(肌肉因子) | IL‑6, irisin, myostatin, IL‑15, BDNF, FGF21, BAIBA, METRNL, myonectin, SPARC, decorin, apelin | 改善胰岛素敏感性、脂质氧化、脂肪棕色化、炎症消退和组织修复 | 机制研究、人类急性运动研究、RCT/对照研究 [1:1][2:1][3:1][7][8][9][10][11][12][13][14][15][16][17] |
| 纤维类型和神经肌肉的维持 | 抵消 II 型纤维的优先萎缩/去神经支配 | 维持爆发力,降低跌倒风险,支持功能独立性 | 组织学、纵向生理学、干预试验 [4:2][18][5:2] |
¶ 1) Muscle mass and autonomy
肌少症(Sarcopenia)是指随着年龄增长,骨骼肌质量、力量和功能进行性丧失;它是残疾、跌倒和死亡率的预测指标 [1:2][4:3]。在数量上,力量的下降速度快于质量的下降(质量变化),例如,纵向队列研究报告称,随着时间的推移,力量和肌肉质量会出现显著流失 [4:4]。
老年人和高龄老人的抗阻训练能在力量和纤维横截面积(CSA)上产生巨大的、具有临床意义的增长,这表明即使在衰弱状态下也是可逆的 [5:3][6:2]。
最低数值(说明性):
- Health ABC 研究:尽管肌肉质量流失不大,但在随访期间力量和肌肉质量显著下降 [4:5]。
- RCT(高龄老人,平均约 87 岁):在进行 8-10 周的高强度抗阻训练后,下肢力量增加约 2-3 倍 [6:3]。
¶ 2) Muscles are glands: skeletal muscle as an endocrine organ
骨骼肌在收缩期间和收缩后会分泌细胞因子/肽类(肌肉因子,myokines)进入血液循环,以自分泌/旁分泌/内分泌的方式发挥作用,调节糖脂代谢、脂肪组织棕色化、炎症消退、组织修复以及大脑功能 [1:3][2:2][3:2][7:1]。
¶ 主要肌肉因子(精选)及其主要作用
| 肌肉因子(首字母缩写展开) | 主要作用 | 典型情境 | 证据 |
|---|---|---|---|
| 白细胞介素-6 (IL‑6) | 增加肌肉中的葡萄糖摄取和脂肪酸氧化;促进脂解;在运动期间急性发挥抗炎信号作用 | 急性运动 → 短暂升高 | 机制和人类运动研究 [1:4][12:1] |
| 鸢尾素 (Irisin)(由 FNDC5 裂解) | 诱导白色脂肪组织褐变;增加产热和能量消耗 | 耐力/抗阻运动 | Nature 发现;人类观察/干预数据 [8:1][13:1] |
| 肌肉生长抑制素 (Myostatin / GDF‑8) | 肌肉生长的负调节因子;抑制 → 肥大 | 基础调节;随训练减少 | 遗传/机制;转化相关性 [19] |
| 白细胞介素-15 (IL‑15) | 支持肌肉合成代谢;与较低的肥胖率相关 | 运动响应 | 人类/生物学研究 [7:2][16:1] |
| 脑源性神经营养因子 (BDNF) | 通过 AMPK 增加脂肪氧化;肌肉-大脑轴(神经营养) | 肌肉收缩;耐力运动 | 机制和人类细胞研究 [9:1][7:3] |
| 成纤维细胞生长因子-21 (FGF21) | 改善胰岛素敏感性;调节葡萄糖/脂质代谢 | 急性和慢性运动;代谢应激 | 人类/小鼠运动/生理学 [14:1][20] |
| β-氨基异丁酸 (BAIBA) | 促进肝脏 β-氧化;诱导白色脂肪褐变 | 运动诱导代谢物 | 人类关联;机制数据 [10:1] |
| 陨石素样蛋白 (METRNL) | 增强米色脂肪产热;免疫-脂肪相互作用 | 寒冷/运动 | 具有转化相关性的细胞和动物研究 [11:1] |
| 肌肉脂联素 (Myonectin / CTRP15) | 增加脂肪酸摄取;激活 mTOR;抑制肝脏自噬 | 运动/肌肉收缩 | 机制和体内研究 [15:1][16:2] |
| 富含半胱氨酸的酸性分泌蛋白 (SPARC) | 代谢作用;结肠中与运动相关的抗肿瘤信号传导 | 规律运动 | 人体组织和动物机制数据 [17:1] |
| 核心蛋白聚糖 (Decorin) | 结合/抑制肌肉生长抑制素;有助于肌肉肥大 | 抗阻运动 | 人类/动物机制研究 [21] |
| 脂肪因子 (Apelin) | 改善肌肉功能/再生;具有良好的代谢作用 | 有氧训练;衰老肌肉 | 人类队列/干预数据 [17:2] |
注:运动期间急性 IL-6 升高与慢性低度炎症不同;情境(时间/组织)决定了净效应 [1:5][12:2]。证据涵盖机制研究、人类急性运动研究和对照训练研究;并非所有肌肉因子都已有明确的临床结局试验 [2:3][7:4][22][23]。
¶ 3) 衰老导致的 II 型(快肌)纤维流失

示意图展示了在人类一生中,与 I 型(慢肌)纤维相比,II 型(快肌)肌肉纤维的优先流失。虽然 I 型纤维保持相对稳定,但 II 型纤维从中年开始经历显著的萎缩和流失。根据 Lexell 等人的数据建立模型。[18:1]
人体骨骼肌包含两种主要的纤维类型:I 型(慢肌),具有抗疲劳性,用于耐力;以及 II 型(快肌),产生高力量和爆发力,但容易疲劳。衰老对这些纤维的影响并不均等。
¶ 优先萎缩与流失
与衰老相关的肌肉流失主要是 II 型纤维的流失。
- 选择性萎缩:II 型纤维在尺寸(横截面积)上显著缩小,在 20 岁至 80 岁之间减少 25–40% 或更多 [18:2][24]。相比之下,I 型纤维通常保持其尺寸。
- 运动单位流失:支配 II 型纤维的 α 运动神经元容易发生凋亡(细胞死亡)。随着这些神经元的死亡,相关的肌肉纤维会失去与神经系统的连接 [18:3]。
¶ “成组”机制
当 II 型纤维失去神经支配时,它会发出求救信号。通常,附近的 I 型运动神经元会长出侧支神经,重新支配这个“孤立”的纤维。这挽救了纤维免于死亡,但代价是:该纤维从 II 型转变为 I 型 [5:4][24:1]。
- 结果:肌肉变得更慢、爆发力更弱。
- 聚集结构:与年轻时混合纤维类型的健康“棋盘”模式不同,衰老的肌肉表现出纤维类型的“成组”现象,表明了这种失神经支配和神经再支配的历史 [18:4]。
¶ 临床后果:爆发力与力量
由于 II 型纤维驱动高速运动,肌肉的爆发力(快速施加力量的能力)比纯粹的力量下降得快得多 [24:2]。这对独立生活能力具有关键影响:
- 跌倒风险:从绊倒中恢复需要瞬间的爆发性迈步(II 型活动)。
- 活动能力:从椅子上站起或爬楼梯需要爆发力。
- 解决方案:抗阻训练,特别是意在快速移动重物的训练(爆发力训练),可以使剩余的 II 型纤维肥大,并提高运动单位的放电频率,从而部分逆转这种衰退 [5:5][6:4]。
¶ 实际意义(基于证据)

一张说明运动诱导的肌肉因子对不同身体系统产生全身益处的信息图。
- 优先进行抗阻训练(每周 2–3 次):增加老年人(包括高龄/体弱者)的力量和 II 型肌纤维 CSA [5:6][6:5]。
- 结合有氧运动:支持肌肉因子谱(IL‑6、FGF21、irisin、apelin)、线粒体功能和心脏代谢健康 [1:6][2:4][7:5][14:2][17:3][20:1]。
- 循序渐进;监测运动强度和恢复情况;结合充足的蛋白质以支持肌肉肥大(此处不作展开)。
¶ 参考文献
Pedersen BK, Febbraio MA. Muscle as an endocrine organ: focus on muscle‑derived interleukin‑6. Physiol Rev. 2008;88(4):1379‑1406. https://journals.physiology.org/doi/full/10.1152/physrev.90100.2007 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Severinsen MCK, Pedersen BK. Muscle–organ crosstalk: the emerging roles of myokines. Endocr Rev. 2020;41(4):594‑609. https://pubmed.ncbi.nlm.nih.gov/32374815/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Laurens C, Bergouignan A, Moro C. Exercise‑Released Myokines in the Control of Energy Metabolism. Front Physiol. 2020;11:91. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7031345/ ↩︎ ↩︎ ↩︎
Goodpaster BH, et al. Loss of strength, mass, and quality in older adults (Health ABC). J Gerontol A. 2006;61(10):1059‑1064. https://academic.oup.com/biomedgerontology/article/61/10/1059/545851 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Frontera WR, Meredith CN, O’Reilly KP, Knuttgen HG, Evans WJ. Strength conditioning in older men: hypertrophy and function. J Appl Physiol. 1988;64(3):1038‑1044. https://journals.physiology.org/doi/abs/10.1152/jappl.1988.64.3.1038 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Fiatarone MA, O’Neill EF, Ryan ND, et al. Exercise training in very elderly people (RCT). N Engl J Med. 1994;330(25):1769‑1775. https://www.nejm.org/doi/full/10.1056/NEJM199406233302501 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Green DJ, et al. The Potential Role of Contraction‑Induced Myokines in Metabolic Regulation. Front Endocrinol (Lausanne). 2017;8:97. https://www.frontiersin.org/articles/10.3389/fendo.2017.00097/full ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Boström P, et al. A PGC1‑α‑dependent myokine (irisin) that drives browning of white fat. Nature. 2012;481:463‑468. https://www.nature.com/articles/nature10777 ↩︎ ↩︎
Wrann CD, et al. Exercise induces hippocampal BDNF via a PGC‑1α/FNDC5 pathway. Cell Metab. 2013;18(5):649‑659. https://www.cell.com/cell-metabolism/fulltext/S1550-4131(13)00372-3 ↩︎ ↩︎
Roberts LD, et al. β‑Aminoisobutyric acid induces browning of white fat and hepatic β‑oxidation. Cell Metab. 2014;19(1):96‑108. https://www.cell.com/cell-metabolism/fulltext/S1550-4131(13)00478-0 ↩︎ ↩︎
Rao RR, et al. Meteorin‑like regulates beige fat thermogenesis. Cell. 2014;157(6):1279‑1291. https://www.cell.com/fulltext/S0092-8674(14)00578-3 ↩︎ ↩︎
Akerström T, et al. Exercise induces IL‑6 release from human skeletal muscle; role in lipid metabolism. Am J Physiol Endocrinol Metab. 2005;288:E733‑E740. https://journals.physiology.org/doi/full/10.1152/ajpendo.00340.2004 ↩︎ ↩︎ ↩︎
Huh JY, et al. Irisin stimulates muscle growth‑related genes and regulates adipocyte metabolism in humans. Int J Obes (Lond). 2014;38:1538‑1544. https://www.nature.com/articles/ijo201442 ↩︎ ↩︎
Izumiya Y, et al. FGF21 is an Akt‑regulated myokine. FEBS Lett. 2008;582(27):3805‑3810. https://febs.onlinelibrary.wiley.com/doi/full/10.1016/j.febslet.2008.10.021 ↩︎ ↩︎ ↩︎
Seldin MM, et al. Myonectin (CTRP15) links skeletal muscle to systemic lipid homeostasis. J Biol Chem. 2012;287(15):11968‑11980. https://www.jbc.org/article/S0021-9258(20)50094-2/fulltext ↩︎ ↩︎
Seldin MM, et al. Skeletal muscle‑derived myonectin activates mTOR to suppress hepatic autophagy. J Biol Chem. 2013;288(50):36073‑36082. https://www.jbc.org/article/S0021-9258(20)50092-3/fulltext ↩︎ ↩︎ ↩︎
Aoi W, Naito Y, Takagi T, et al. SPARC and exercise‑linked suppression of colon tumorigenesis. Gut. 2013;62(6):882‑889. https://gut.bmj.com/content/62/6/882.long ↩︎ ↩︎ ↩︎ ↩︎
Lexell J, Taylor CC, Sjöström M. Ageing atrophy: number/size/proportion of fiber types in vastus lateralis (15–83 y). J Neurol Sci. 1988;84:275‑294. https://www.sciencedirect.com/science/article/abs/pii/0022510X88901245 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
McPherron AC, Lawler AM, Lee SJ. Regulation of skeletal muscle mass by a new TGF‑β family member (myostatin). Nature. 1997;387:83‑90. https://www.nature.com/articles/387083a0 ↩︎
Kim KH, Lee MS, et al. Acute exercise induces FGF21 in mice and humans. PLoS One. 2013;8(5):e63517. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0063517 ↩︎ ↩︎
Kanzleiter T, Rath M, Görgens SW, et al. The myokine decorin is regulated by contraction and involved in hypertrophy. Biochem Biophys Res Commun. 2014;450(2):1089‑1094. https://www.sciencedirect.com/science/article/pii/S0006291X14011089 ↩︎
Hoffmann C, Weigert C. Skeletal muscle as an endocrine organ: myokines in exercise adaptations. Cold Spring Harb Perspect Med. 2017;7(11):a029793. https://pubmed.ncbi.nlm.nih.gov/28389517/ ↩︎
Pedersen BK, Febbraio MA. Muscles, exercise and obesity: skeletal muscle as a secretory organ. Nat Rev Endocrinol. 2012;8(8):457‑465. https://www.nature.com/articles/nrendo.2012.49 ↩︎
Verdijk LB, et al. Skeletal muscle hypertrophy following resistance training is accompanied by a fiber type-specific increase in satellite cell content in elderly men. J Gerontol A Biol Sci Med Sci. 2009;64(3):332-339. https://academic.oup.com/biomedgerontology/article/64A/3/332/625396 ↩︎ ↩︎ ↩︎