¶ NAD+ (烟酰胺腺嘌呤二核苷酸)
¶ 概览
定义:NAD+ 是一种存在于所有活细胞中的通用电子载体和酶促辅因子。它是“代谢货币”,对于将营养物质转化为能量(ATP)至关重要,同时也是长寿蛋白消耗的燃料。
衰老危机:从青年期到老年期,NAD+ 水平会下降高达 50%。这种水平的崩溃是线粒体功能障碍、基因组不稳定和细胞衰老的主要驱动因素。[1]
生物学作用:
- 能量:如果没有 NAD+,线粒体的能量产生会立即停止。
- 信号传导:它是 Sirtuins(长寿基因)和 DNA 修复酶的“启动开关”。
干预策略:NAD+ 本身的口服生物利用度很差,且不易穿过细胞膜。临床策略主要集中在细胞可以转化为 NAD+ 的前体(如 NMN 和 NR)上。
👉 有关干预方案、剂量和补充剂的信息,请参阅 NAD+ 前体。
¶ 为什么重要:生命的燃料
NAD+ 以两种截然不同的形式存在,它们之间的比例决定了细胞的代谢状态。
¶ 1. 氧化还原引擎 (NADH)
在其主要作用中,NAD+ 充当电子穿梭机。它接受来自葡萄糖和脂肪酸分解的高能电子,变成 NADH(还原形式)。然后,它将这些电子提供给线粒体中的电子传递链(Electron Transport Chain)以生成 ATP。
- 循环:在年轻、健康的细胞中,NAD+ 和 NADH 之间不断地来回循环。这起到了催化剂的作用——它不会被消耗,只是被重复利用。
¶ 2. 消耗性信号分子
在其对衰老至关重要的次要作用中,NAD+ 会被酶永久消耗。它被分解以为修复 DNA 和调节基因表达的反应提供能量。
- 冲突:由于这些酶会破坏 NAD+,它们会与线粒体争夺有限的 NAD+ 库。如果消耗量超过了细胞制造更多 NAD+(合成)的能力,整个系统就会崩溃。
¶ 作用机制:长寿通路
NAD+ 是参与衰老标志(Hallmarks of Aging)的三大类酶的专性底物。
¶ 1. Sirtuins:“基因组的守护者”
Sirtuins(SIRT1–SIRT7)是一个需要 NAD+ 才能发挥作用的脱酰酶家族。没有它,它们就无法运作。
- SIRT1:调节昼夜节律、线粒体生物发生(通过 PGC-1α),并沉默“垃圾”基因。[2]
- SIRT3:定位于线粒体,维持抗氧化防御(SOD2)并确保高效的 ATP 产生。
- SIRT6:对 DNA 双链断裂修复和维持端粒稳定性至关重要。
- 与衰老的联系:Sirtuins 对 NAD+ 的亲和力较低(高 ),这意味着当 NAD+ 水平下降时,它们是最先关闭的。恢复 NAD+ 可以“唤醒”这些休眠的防御系统。
¶ 2. PARPs:DNA 修复小队
聚 ADP-核糖聚合酶(PARPs),特别是 PARP1,是 DNA 损伤的“第一响应者”。
- 机制:当 PARP1 检测到 DNA 链断裂时,它会结合到损伤处,并利用 NAD+ 分子构建长链(聚 ADP-核糖),向其他修复酶发出信号以修复断裂。
- 代价:修复单个 DNA 断裂可能会消耗大量的 NAD+。
¶ 3. CD38:免疫调节剂
CD38 是一种膜结合酶,存在于免疫细胞(巨噬细胞、B 细胞)上,并在衰老过程中广泛存在于组织中。
- 机制:它降解 NAD+ 以产生环 ADP-核糖(cADPR),这是一种用于钙信号传导的分子。
- 问题:CD38 是一种效率极低的酶——它每产生 1 个分子的 cADPR 信号,就会消耗大约 100 个分子的 NAD+,充当了一个巨大的“消耗漏斗”。[3]
¶ 衰老问题:为什么 NAD+ 会下降
与年龄相关的 NAD+ 崩溃并不是一个简单的生产问题;这是一场由两股汇聚的力量引起的“供需”危机:消耗增加和生产减少。
¶ 1. “消耗漏斗”:慢性炎症与 CD38
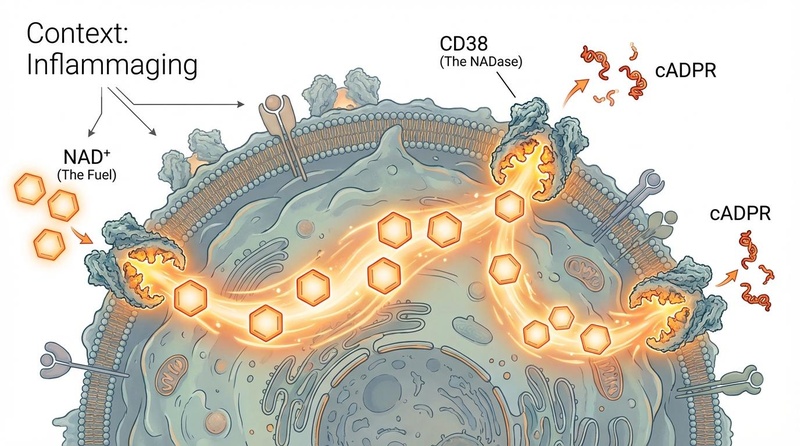
在衰老组织中,NAD+ 下降的最重要驱动因素是 CD38 的上调。
- 炎性衰老 (Inflammaging):随着年龄的增长,不断积累的衰老细胞会分泌炎症因子 (SASP),例如 TNF-α 和 IL-6。
- 恶性循环 (The Viscous Cycle):这些炎症细胞因子触发巨噬细胞(免疫细胞)增殖并表达大量的 CD38。这产生了一个系统性的“NAD+ 消耗漏斗”,不仅消耗免疫细胞中的 NAD+,还消耗周围健康组织中的 NAD+。[4]
- 证据:在老年小鼠中,脂肪和肝脏组织中的 CD38 水平可能比年轻小鼠高 2-3 倍。[5]
¶ 2. “燃烧殆尽”:DNA 损伤与 PARPs
衰老伴随着来自紫外线辐射、氧化应激和复制错误的 DNA 损伤的积累。
- 慢性激活:这种持续的低水平损伤使 PARP 酶保持慢性活跃状态。它们不断从 NAD+ 池中提取能量来为修复提供动力。
- 过度激活:在严重损伤的情况下,PARP1 会将细胞内的 NAD+ 消耗至接近零的水平,导致能量衰竭和细胞死亡(程序性坏死,necroptosis)。[6]
¶ 3. “瓶颈”:NAMPT 下降
在消耗量激增的同时,身体回收 NAD+ 的能力却在衰退。
- NAMPT:补救途径(salvage pathway,见下文)中的这种限速酶会随着年龄和代谢应激而下降。[7]
- 昼夜节律紊乱:NAMPT 的表达受昼夜节律钟(BMAL1/CLOCK)的调节。与年龄相关的睡眠碎片化和昼夜节律失调会抑制 NAMPT,进一步阻断供应。[8]
¶ 生物合成:身体如何制造 NAD+
哺乳动物依赖三种途径来维持 NAD+ 池。在衰老过程中,这种机制从高效的回收利用转变为对前体的迫切需求。
¶ 1. 补救途径(主导)
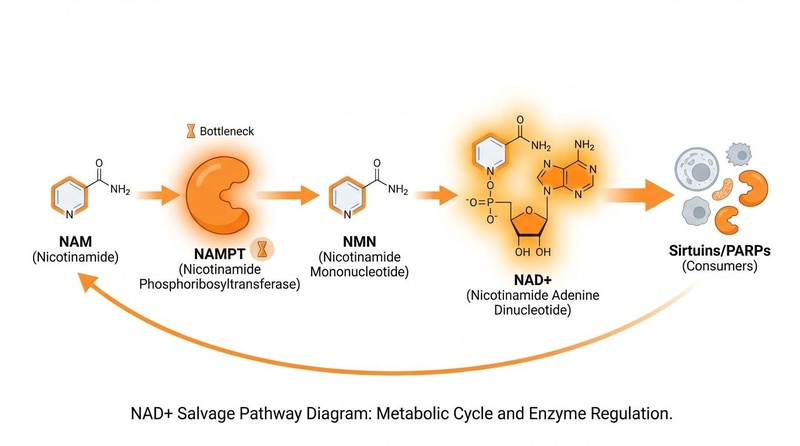
该途径将 NAD+ 使用后的“废物”产物——烟酰胺(Nicotinamide,NAM)——回收成新的 NAD+。
- 途径:NAM → NMN(通过 NAMPT)→ NAD+。
- 作用:它为大多数组织提供了绝大部分的 NAD+。
- 脆弱性:由于它依赖于 NAMPT,该途径最容易受到衰老和炎症的破坏。
¶ 2. Preiss-Handler 途径
该途径利用饮食中的烟酸(Nicotinic Acid,即 Niacin/维生素 B3)。
- 途径:烟酸(NA)→ NaMN → NaAD → NAD+。
- 作用:它绕过了 NAMPT 瓶颈,但受到高剂量烟酸的“潮红”副作用及其酶的组织特异性表达的限制。[9]
¶ 3. 从头合成途径(犬尿氨酸)
该途径利用氨基酸色氨酸(Tryptophan)从头合成 NAD+。
- 途径:色氨酸 → ...(多步反应)... → NAD+。
- 效率:极低。大约需要 60 毫克色氨酸才能产生 1 毫克烟酸当量。它在能量上消耗巨大,无法支持衰老身体的高需求。[10]
¶ 衰退机制总结
| 机制 | 原因 | 对 NAD+ 的影响 |
|---|---|---|
| CD38 上调 | 慢性炎症(SASP) | 大量消耗(主要消耗源) |
| PARP 激活 | 积累的 DNA 损伤 | 高消耗(可变消耗源) |
| NAMPT 下调 | 衰老、昼夜节律紊乱 | 回收减少(供应衰竭) |
临床意义:有效恢复 NAD+ 可能需要解决等式的两边:提供前体(以绕过 NAMPT)和减少炎症(以降低 CD38)。
👉 有关恢复 NAD+ 的可操作策略,请参阅 NAD+ 前体。
¶ 参考文献
Covarrubias, A. J., Perrone, R., Grozio, A., & Verdin, E. (2021). NAD+ metabolism and its roles in cellular processes during ageing. Nature Reviews Molecular Cell Biology, 22(2), 119-141. https://www.nature.com/articles/s41580-020-00313-x ↩︎
Imai, S., & Guarente, L. (2014). NAD+ and sirtuins in aging and disease. Trends in Cell Biology, 24(8), 464-471. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4112140/ ↩︎
Chini, E. N. (2009). CD38 as a regulator of cellular NAD: a novel potential pharmacological target for metabolic conditions. Current Pharmaceutical Design, 15(1), 57-63. https://pubmed.ncbi.nlm.nih.gov/19149601/ ↩︎
Chini, C. C., et al. (2020). CD38 ecto-enzyme in immune cells is induced during aging and regulates NAD+ and NMN levels. Nature Metabolism, 2, 1284–1304. https://www.nature.com/articles/s42255-020-00298-z ↩︎
Camacho-Pereira, J., et al. (2016). CD38 Dictates Age-Related NAD Decline and Mitochondrial Dysfunction through an SIRT3-Dependent Mechanism. Cell Metabolism, 23(6), 1127-1139. https://www.cell.com/cell-metabolism/fulltext/S1550-4131(16)30224-8 ↩︎
Pacher, P., & Szabo, C. (2008). Role of the peroxynitrite-poly(ADP-ribose) polymerase pathway in human disease. American Journal of Pathology, 173(1), 2-13. https://pmc.ncbi.nlm.nih.gov/articles/PMC2438291/ ↩︎
Yoshino, J., Mills, K. F., Yoon, M. J., & Imai, S. (2011). Nicotinamide mononucleotide, a key NAD+ intermediate, treats the pathophysiology of diet- and age-induced diabetes in mice. Cell Metabolism, 14(4), 528-536. https://www.cell.com/cell-metabolism/fulltext/S1550-4131(11)00345-8 ↩︎
Ramsey, K. M., et al. (2009). Circadian clock feedback cycle through NAMPT-mediated NAD+ biosynthesis. Science, 324(5927), 651-654. https://pubmed.ncbi.nlm.nih.gov/19342540/ ↩︎
Bogan, K. L., & Brenner, C. (2008). Nicotinic acid, nicotinamide, and nicotinamide riboside: a molecular evaluation of NAD+ precursor vitamins in human nutrition. Annual Review of Nutrition, 28, 115-130. https://pubmed.ncbi.nlm.nih.gov/18429699/ ↩︎
Badawy, A. A. (2017). Kynurenine Pathway of Tryptophan Metabolism: Regulatory and Functional Aspects. International Journal of Tryptophan Research, 10. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5398323/ ↩︎