¶ MicroRNAs
MicroRNAs (miRNAs) represent a highly conserved class of endogenous, short (typically 19–25 nucleotides), non-coding RNA molecules that exert post-transcriptional control over gene expression. By hybridizing to complementary sequences within the 3' untranslated regions (UTRs) of target messenger RNAs (mRNAs), miRNAs initiate either translational repression or enzymatic cleavage, effectively "fine-tuning" the cellular proteome.
In the context of biogerontology and clinical longevity medicine, microRNAs serve a critical dual role. First, they act as highly stable, biofluid-accessible biomarkers ("senomiRs" and "inflamma-miRs") that reflect systemic aging trajectories, organ-specific dysfunction, and senescent-cell burden [1]. Second, they function as therapeutic targets and interventions, where synthetic miRNA mimics or antisense inhibitors (antagomirs) are deployed to reprogram pathological pathways and restore youthful cellular homeostasis.
| Indication | Research / Off-label (Longevity pathways, age-related diseases) |
| Access | Investigational / Research Only / Phase II & III Trials |
| Dosing Sched | Delivery-dependent (e.g., weekly subcutaneous, local intradermal, or daily oral small molecules) |
| Safety Profile | Moderate (Dependent on vector specificity, immunogenicity, and off-target risks) |
| Key Marker | Circulating miRNA profile, target gene transcription levels, systemic inflammatory panels |
| Est. Cost | High (Investigational clinical-grade therapies) |
¶ At a Glance
MicroRNAs function as post-transcriptional regulators, serving as pivotal nodes in longevity networks. Their capacity to simultaneously modulate multiple downstream genes makes them exceptionally powerful—yet highly complex—clinical targets.
¶ Key Points
- Systemic Regulation: A single microRNA can coordinate entire genetic networks, influencing hallmarks of aging such as cellular senescence, chronic inflammation, and tissue regeneration [2].
- Diagnostic Precision: Extracellular miRNAs packaged in exosomes are exceptionally stable in circulation, providing non-invasive diagnostic readouts of biological age and cellular senescence cell burden [3][4].
- Delivery Bottlenecks: The clinical translation of miRNA therapeutics is heavily constrained by delivery vector safety, as first-generation lipid nanoparticles (LNPs) can trigger severe immune reactions, and uncontrolled viral delivery can lead to aberrant tissue growth [5][6].
¶ What People Use It For
- Primary Objectives: Systemic senescence clearance, myocardial and cutaneous tissue regeneration, suppression of chronic vascular inflammation, and diagnostic profiling of biological aging.
- Evidence Quality: High for localized interventions (fibrosis, viral targets) and observational biomarkers; Moderate to Low for systemic longevity therapies in early-stage clinical development.
¶ What is it?
MicroRNAs are endogenous RNA molecules that act as the master "dimmer switches" of the mammalian genome. Discovered to be pivotal in developmental timing and cellular differentiation, they do not encode proteins but instead govern the stability and translation of protein-coding mRNAs [2:1].
Unlike classic antisense oligonucleotides (ASOs) that target a single specific transcript, a single miRNA typically possesses a "seed sequence" (nucleotides 2–8) that matches consensus elements on dozens or even hundreds of distinct mRNAs. This multi-target capability allows miRNAs to act as systems-level regulators, coordinating complex physiological responses such as cellular stress defense, metabolic remodeling, and inflammatory signaling cascades.
Core Therapeutic Modalities
- miRNA Mimics: Chemically synthesized double-stranded RNA molecules designed to replace or bolster a specific protective miRNA that has declined during biological aging (e.g., miR-29 for anti-fibrosis [7] or miR-34a in oncogene suppression [5:1]).
- Antagomirs (Anti-miRs): Single-stranded, chemically modified oligonucleotides designed to bind with high affinity to endogenous, disease-promoting or pro-senescent miRNAs, neutralizing them and preventing them from silencing protective target mRNAs (e.g., miravirsen targeting hepatic miR-122 [8] or cobomarsen targeting pro-inflammatory miR-155 [9]).
- Indirect Modulators: Small molecule therapeutics that selectively upregulate or downregulate specific miRNA transcription or processing in target tissues, such as obefazimod, which selectively enhances the anti-inflammatory regulator miR-124 [10].
¶ Main Benefits
Targeting microRNAs offers unparalleled advantages in therapeutic design due to their ability to regulate integrated cellular networks rather than single isolated receptors.
¶ Systemic Longevity Signature Profiling
Epidemiological and cohort studies have demonstrated that centenarians and long-lived individuals harbor distinct circulating microRNA profiles. For instance, nonagenarians and centenarians from Okinawan cohorts exhibit a protective "longevity signature" characterized by the sustained upregulation of anti-inflammatory miRNAs (such as miR-181a-5p) and the downregulation of pro-inflammatory, pro-senescent miRNAs (such as miR-21-5p) [11]. Measuring these panels allows clinicians to map patient trajectories along healthy versus unhealthy biological aging pathways [12][4:1]. Circulating miRNAs have also been validated as highly feasible biomarkers of chronological and biological age in older cohorts [13].
¶ Senescence and Fibrosis Reversal
As tissues age, the accumulation of senescent cells and the progressive deposition of collagenous extracellular matrix lead to parenchymal fibrosis and organ stiffening. miRNA-based therapeutics have demonstrated the ability to reverse these changes in clinical models:
- Tissue Remodeling: Intradermal delivery of miR-29 mimics (remlarsen) directly represses the expression of collagen-1, collagen-3, and other extracellular matrix proteins, restoring youthful tissue compliance and preventing pathologic scar formation [7:1].
- SenomiR Diagnostics: Panels of 22 circulating miRNAs (senomiRs) can accurately quantify the systemic cellular senescence cell burden, providing a direct molecular readout of senolytic drug efficacy [3:1].
¶ Anti-Inflammatory Reprogramming (Inflamma-miRs)
Chronic, low-grade, sterile inflammation ("inflammaging") is mediated by the progressive upregulation of pro-inflammatory miRNAs ("inflamma-miRs") [14]. Systemic antagomirs can arrest this cascade. For example, neutralizing miR-155 via cobomarsen suppresses pathogenic T-cell activation and downregulates systemic pro-inflammatory cytokines, presenting a therapeutic avenue for chronic inflammation [9:1]. Similarly, upregulating the cellular "brake" miR-124 using the small molecule obefazimod drives significant and durable reductions in inflammatory cytokines (e.g., IL-1β, IL-6, TNF-α), leading to deep clinical and endoscopic remission in clinical trials [10:1]. Upregulation of miR-124 via obefazimod has also demonstrated high efficacy, anti-inflammatory tissue remodeling, and target engagement in patients with active rheumatoid arthritis [15].
¶ Evidence Summary Table (Human Outcomes)
| Outcome / Goal | Effect* | Consistency** | Evidence Quality | Trials*** | Notes (population, duration, dose) |
|---|---|---|---|---|---|
| Hepatitis C Viral Suppression | High | High | 1 Phase 2a RCT | Miravirsen (anti-miR-122) subcutaneous weekly dosing over 29 days significantly and durably reduced viral load in chronic HCV patients [8:1]. | |
| Ulcerative Colitis Remission | High | High | 1 Phase 2b RCT, Phase 3 | Obefazimod (ABX464 small molecule miR-124 upregulator) once-daily oral dosing significantly reduced modified Mayo score and induced mucosal healing at 8 weeks [10:2]. | |
| Rheumatoid Arthritis Control | High | High | 1 Phase 2 RCT | Obefazimod (ABX464) once-daily oral dosing significantly reduced disease activity and inflammatory cytokine production at 12 weeks [15:1]. | |
| Cutaneous Fibrosis & Scar Suppression | High | High | 1 Phase 1 RCT | Remlarsen (miR-29 mimic) intradermal injections at excisional wound sites safely downregulate collagen and ECM [7:2]. | |
| T-Cell Lymphoma Stabilization | Moderate | Moderate | 1 Phase 1 trial | Cobomarsen (anti-miR-155 LNA antagomir) delivered systemically led to target engagement and disease stabilization [9:2]. | |
| Angiogenesis & Tissue Repair | High | Moderate | 1 Phase 1 trial | MRG-110 (anti-miR-92a LNA ASO) intradermal injection accelerated perfusion, CD49e expression, and blood vessel growth [16][17]. | |
| Tumor Stabilization (Mesothelioma) | Moderate | Moderate | 1 Phase 1 trial | TargomiRs (miR-16 mimic via EGFR-targeted minicells) yielded stable disease in 68% of mesothelioma patients [18]. | |
| Solid Tumor Suppression (Systemic LNP) | Low | Low | 1 Phase 1 trial | MRX34 (miR-34a mimic via LNPs) trial halted due to severe immune-mediated adverse events and deaths [5:2]. | |
| All-Cause Mortality Risk Prediction | High | Moderate | Cohort Study | miRNA-based biological age models (mirAge, mirMort, mirFI) predict self-reported health decline, frailty, and 10-year all-cause mortality [4:2]. | |
| Extreme Longevity Biomarkers | High | Moderate | Observational | Centenarian serum profiles show protective up-regulation of miR-181a-5p and downregulation of inflammatory miR-21-5p [11:1]. | |
| Systemic Senescence Burden Diagnostics | Moderate | Moderate | Cohort Study | Panel of 22 circulating senomiRs (including miR-31-5p, miR-19a-3p) stably increases with age and correlates with tissue senescent-cell accumulation [3:2][19]. |
- *Effect: Compact renderer encoding represents the direction and magnitude of the therapeutic or biomarker outcome.
- **Consistency: Low (conflicting clinical results), Moderate (mixed but leaning positive), High (most trials and studies agree).
- ***Trials: Number of RCTs, clinical trials, or prospective cohorts informing the clinical endpoint.
¶ Mechanisms
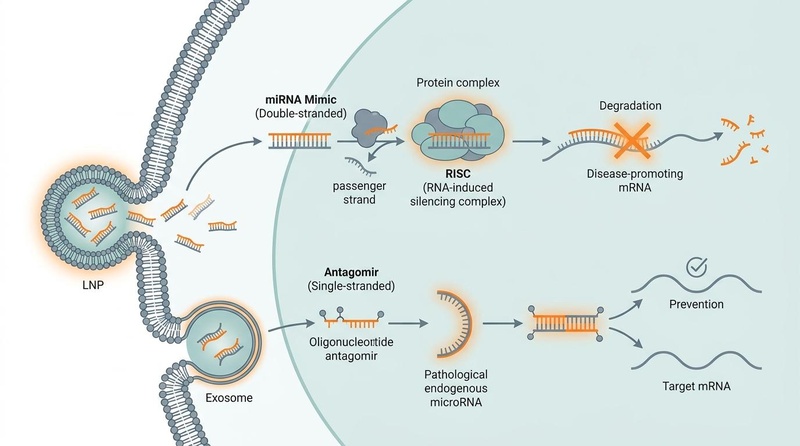
MicroRNAs function via base-pairing with target mRNAs. This molecular mechanism and its clinical manipulation are detailed in the schematic below:
Fig. 1: Clinical mechanisms of microRNA-based therapeutics. On the left, double-stranded synthetic miRNA mimics are delivered via lipid nanoparticles (LNPs), separating into a passenger strand and a guide strand; the guide strand loads into the RNA-induced silencing complex (RISC) to target and degrade disease-promoting mRNA. On the right, single-stranded oligonucleotide antagomirs delivered via exosomes bind and sequester pathological mature endogenous miRNAs, preventing them from silencing target mRNAs.
The intracellular mechanism of action is mediated by the RNA-induced silencing complex (RISC). The primary catalytic engine of RISC is Argonaute (AGO) protein, which recruits the single-stranded miRNA "guide" strand. When the miRNA guide exhibits perfect or near-perfect complementarity to the target mRNA (common in plants and synthetic mimics), RISC induces direct endonucleolytic cleavage of the transcript. More commonly in mammalian systems, partial complementarity (typically in the 3' UTR) results in translational repression and subsequent deadenylation, accelerating mRNA decay [2:2].
¶ Pivotal Longevity and Cellular Pathways
The p53/miR-34a/SIRT1 Senescence Loop
One of the most heavily studied miRNA pathways in mammalian aging is the p53/miR-34a/SIRT1 feed-forward loop. Under chronic cellular stress or DNA damage, the tumor suppressor p53 is activated, directly transcribing the pro-senescent microRNA miR-34a [12:1]. In turn, miR-34a directly binds to and represses the mRNA of Sirtuin 1 (SIRT1)—a critical NAD+-dependent deacetylase that promotes DNA repair, mitochondrial biogenesis, and cell survival [12:2][20].
As SIRT1 levels fall, p53 remains highly acetylated and active, driving further miR-34a transcription and cementing cellular senescence. Therapeutically breaking this loop using antagomirs targeting miR-34a represents a premier strategy to rescue SIRT1 expression, restore mitochondrial function, and rejuvenate senescent stem cell niches [12:3][20:1].
Chronic Stress/DNA Damage ──> p53 Activation
│
▼
miR-34a Transcription ──┐
▲ │ (Inhibition)
│ ▼
p53 Acetylation <── SIRT1 Repression
(Highly Active)
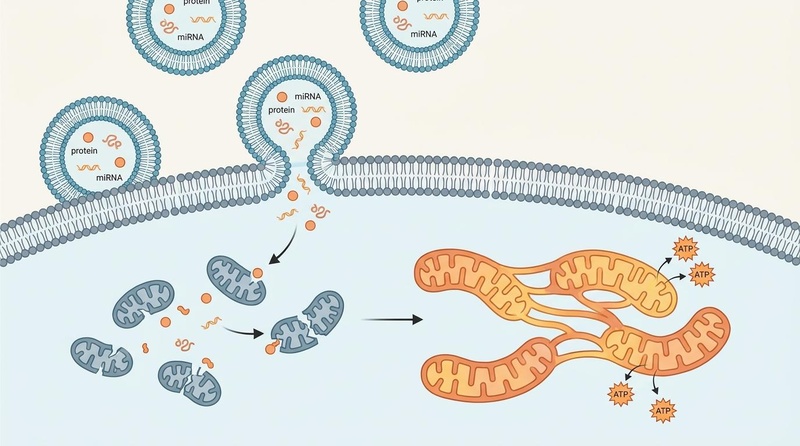
Fig. 2: Exosomal delivery of miRNA promoting mitochondrial network fusion. Following cellular uptake of extracellular vesicles (exosomes), the released miRNA and protein cargo promote the fusion of fragmented, dysfunctional mitochondria into a healthy, interconnected tubular network, restoring cellular ATP production and reversing senescent phenotypes.
Vascular Integrity & Endothelial Function (miR-126)
Vascular aging is driven by progressive endothelial dysfunction. The endothelium is highly dependent on miR-126, which maintains vascular integrity, suppresses leukocyte adhesion, and promotes angiogenesis by targeting inhibitors of the VEGF signaling pathway (specifically SPRED1 and PIK3R2) [21]. During chronological aging and in metabolic diseases such as type 2 diabetes, circulating and tissue levels of miR-126 are dramatically lost, leading to accelerated cardiovascular risk and poor ischemia recovery [21:1][22].
Epigenetic Alterations and the Hallmarks of Aging
Beyond direct translational blockages, miRNAs interact dynamically with the epigenetic landscape. "Epi-miRs" (such as the miR-29 family) directly target DNA methyltransferases (DNMT3A and DNMT3B), thereby modulating global DNA methylation patterns. Loss of miR-29 with age leads to the hypermethylation of genomic loci and the down-regulation of protective tissue-remodeling genes, illustrating how miRNA dysregulation propagates across multiple hallmarks of aging [7:3][2:3].
¶ Dosage & Protocols
Because microRNA therapies are investigational and subject to rapid biological clearance, their clinical application relies heavily on advanced biochemical modifications and targeted delivery vehicles.
¶ Delivery Technologies and Formulations
¶ Lipid Nanoparticles (LNPs)
Commonly used to package nucleic acids inside a protective lipid envelope, LNPs shield miRNAs from enzymatic degradation in circulation and facilitate cell entry via endocytosis [5:3]. However, non-targeted LNPs accumulate predominantly in the liver and spleen, and can trigger severe systemic immune activation. Systemic LNP-mediated delivery of miR-34a mimics (MRX34), for example, was limited by high immunogenicity and toxic systemic cytokine release [5:4].
¶ Exosomes (Extracellular Vesicles)
Naturally occurring nanoscale vesicles (30–150 nm) that transfer biomolecules between cells. Exosomes present membrane proteins (such as CD9, CD63, CD81, and integrins) that facilitate biocompatible, low-immunogenicity delivery across physiological barriers [10:3][19:1]. Because they bypass the severe immune responses triggered by synthetic LNPs, engineered exosomes are considered the gold-standard vehicle for delivering anti-senescent miRNA mimics and antagomirs [19:2].
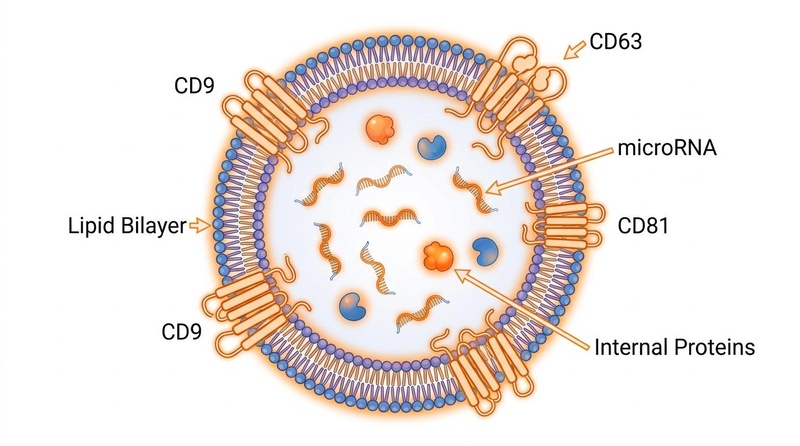
Fig. 3: Structure of an extracellular vesicle (exosome). The vesicle consists of a phospholipid lipid bilayer membrane embedded with tetraspanin proteins (CD9, CD63, and CD81) enclosing an internal cargo of therapeutic proteins and microRNAs.
¶ Targeted Bacterial Minicells (EDVs)
EnGeneIC Dream Vectors (EDVs) are non-viable, bacterially derived minicells coated with cell-specific bispecific antibodies (e.g., anti-EGFR). This design directs the minicell cargo (such as a miR-16 mimic, TargomiRs) straight to target cells, avoiding systemic tissues and preventing liver toxicity [18:1].
¶ Antisense Oligonucleotides (ASOs) & Chemical Modifications
To extend half-life and improve tissue uptake, single-stranded antagomirs are modified with chemistries such as:
- Locked Nucleic Acids (LNAs): Conformationally restricted nucleotides that dramatically increase hybridization stability and thermal melting temperature, allowing for shorter, highly stable antagomirs [8:2][9:3].
- 2'-O-Methyl (2'-O-Me) and 2'-Fluoro (2'-F) modifications: Prevent nuclease cleavage.
- Phosphorothioate (PS) linkages: Prevent rapid clearance by renal filtration.
- Cholesterol Conjugation (Chol-conjugated): Facilitates direct transmembrane transport without requiring a nanoparticle carrier [7:4].
¶ Dosing Paradigms in Clinical Studies
As there is no standardized over-the-counter dosing regimen for microRNAs, clinical and research protocols are indication-specific:
- Localized Fibrosis Control (Remlarsen, miR-29 mimic): Administered via local intradermal injections at a dose of 3.0 mg to 10.0 mg per centimeter of wound margin, repeated weekly during active healing phases to suppress keloid formation [7:5].
- Systemic Inflammatory Control (Obefazimod, miR-124 upregulator): Administered as a once-daily oral dose of 50 mg to 100 mg. This small molecule upregulates miR-124 in immune cells, suppressing inflammatory pathways sustainably over 8–48 weeks [10:4].
- Systemic Antagomir Delivery (Miravirsen, anti-miR-122): Administered via subcutaneous injections at doses ranging from 3.0 to 7.0 mg/kg, delivered weekly for 5 consecutive weeks, providing sustained viral suppression for up to 14 weeks due to the deep tissue half-life of LNA-modified compounds [8:3].
¶ Safety
The safety profile of microRNA therapeutics is highly dependent on the delivery vehicle, the precision of tissue targeting, and the extent of off-target hybridization.
¶ Critical Safety Ceiling: Immunogenicity & Inflammation
Systemic administration of synthetic double-stranded miRNA mimics carries a high risk of triggering the innate immune system. In the Phase I trial of MRX34 (an LNP-encapsulated miR-34a mimic), systemic delivery caused severe, fatal immune-mediated adverse events (including pneumonitis, colitis, and cytokine release syndrome) in multiple patients, resulting in the termination of the trial [5:5]. Double-stranded RNA can activate Toll-like receptors (TLR3, TLR7, TLR8) and cytosolic sensors, initiating a profound inflammatory response.
¶ Spatiotemporal Hazard: Uncontrolled Regeneration
While microRNAs possess remarkable regenerative potential, continuous, unregulated expression can have catastrophic consequences. In a landmark Nature study, delivery of human miR-199a via a viral vector (AAV6) stimulated powerful cardiomyocyte proliferation and scar reduction in pigs following myocardial infarction [6:1]. However, because the viral vector could not be turned off, expression remained active, leading to the formation of poorly differentiated, highly arrhythmogenic myoblasts. This triggered sudden, lethal ventricular tachyarrhythmias in 100% of the treated animals at 7–8 weeks post-treatment, illustrating the absolute necessity for transient, strictly controlled expression systems [6:2].
¶ Off-Target Hybridization
Because miRNAs require only partial complementarity (primarily the 7-nucleotide seed sequence) to bind and repress target transcripts, synthetic mimics or high-dose antagomirs can inadvertently bind and suppress unintended mRNAs in healthy tissues. This off-target silencing can disrupt critical homeostatic pathways, highlighting the need for highly localized delivery or tissue-specific targeting vectors [6:3][2:4].
¶ Therapeutic Discontinuation and Clinical Monitoring Indicators
Investigational protocols utilizing systemic RNA therapeutics require rigid clinical monitoring to detect early markers of toxicity. Indicators for immediate therapeutic cessation include:
- Systemic inflammatory markers rising rapidly above baseline (e.g., C-Reactive Protein > 10 mg/L or acute elevations in IL-6).
- Platelet count dropping below 100,000/μL (indicative of oligonucleotide-induced thrombocytopenia).
- Liver transaminases (ALT/AST) elevating to >3 times the upper limit of normal, indicating hepatic saturation and toxicity.
- Unexplained pulmonary symptoms, dyspnea, or gastrointestinal distress, which may herald early-stage immune-mediated pneumonitis or colitis.
¶ Practical FAQ
What is the difference between an siRNA and a microRNA?
While both are small non-coding RNAs, small interfering RNAs (siRNAs) are typically exogenous in origin, designed with 100% sequence complementarity to target and knock down a single specific mRNA transcript. In contrast, microRNAs are endogenous regulators designed with partial complementarity, enabling them to target and coordinate entire networks of dozens or hundreds of mRNAs simultaneously [2:5].
Can circulating microRNAs be used to measure my biological age?
Yes. Clinical and cohort studies have validated specific panels of circulating microRNAs as highly sensitive aging biomarkers. Biological age models trained on microRNA expression profiles (such as mirAge, mirMort, and mirFI) correlate robustly with physical functioning, frailty, and 10-year all-cause mortality, outperforming chronological age in risk prediction [4:3].
Why can't I buy microRNA supplements online?
Synthetic microRNAs are highly unstable and are rapidly degraded by RNA nucleases in the blood within minutes if not protected by chemical modifications or complex delivery vectors [5:6]. Furthermore, systemic delivery carries high risks of immunogenicity and severe off-target effects. Therefore, microRNA therapies remain restricted to investigational clinical trials under strict monitoring [5:7][6:4].
How do exosomes improve the delivery of microRNA therapeutics?
Exosomes are biocompatible, membrane-bound vesicles produced naturally by human cells. Because they express native surface proteins, they can bypass immune recognition, avoid rapid clearance by the liver, and deliver miRNAs directly into target tissues. This makes them significantly safer and more effective carriers than synthetic lipid nanoparticles (LNPs) [10:5][19:3].
Is there a way to naturally modulate my microRNAs?
Certain lifestyle interventions, such as regular physical exercise and caloric restriction, have been shown in observational studies to partially restore youthful microRNA expression profiles, such as upregulating protective miR-126 and downregulating pro-inflammatory inflamma-miRs, though these changes are modest compared to direct therapeutic interventions [2:6][22:1].
¶ References
Ibáñez-Ventoso C, et al. Can microRNAs act as biomarkers of aging? Aging (Albany NY). 2010;2(11):736-747. https://ncbi.nlm.nih.gov/pmc/articles/PMC3044429 ↩︎
Kinser HE, Pincus Z. MicroRNAs as modulators of longevity and the aging process. Hum Genet. 2020;139(1):17-31. https://pmc.ncbi.nlm.nih.gov/articles/PMC6954352/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Grillari J, et al. Profiling microRNA expression during senescence and aging: mining for a diagnostic tool of senescent-cell burden. bioRxiv. 2024;2024.04.10.588794. https://www.biorxiv.org/content/10.1101/2024.04.10.588794v2 ↩︎ ↩︎ ↩︎
Rotterdam Study Consortium. Plasma microRNA signatures of aging and their links to health outcomes and mortality: findings from a population-based cohort study. Genome Med. 2025;17(1):15. https://pmc.ncbi.nlm.nih.gov/articles/PMC12188677/ ↩︎ ↩︎ ↩︎ ↩︎
Beg MS, Brenner AJ, Sachdev J, et al. Phase I Study of MRX34, a Liposome-Formulated mR-34a Mimic, in Patients With Advanced Solid Tumors. J Clin Oncol. 2019;37(14):1128-1136. https://ascopubs.org/doi/full/10.1200/JCO.18.01254 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Gabisonia K, Prosdocimo G, Diez-Ciudad I, et al. MicroRNA therapy stimulates uncontrolled cardiac repair after myocardial infarction in pigs. Nature. 2019;575(7782):356-362. https://www.nature.com/articles/s41586-019-1191-6 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Gallant-Behm CL, Zhang Y, Lee SJ, et al. A MicroRNA-29 Mimic (Remlarsen) Represses Extracellular Matrix Expression and Fibroplasia in the Skin. J Invest Dermatol. 2019;139(3):614-623. https://www.sciencedirect.com/science/article/pii/S0022202X18328306 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Janssen HL, Reesink HW, Lawitz EJ, et al. Treatment of HCV Infection by Targeting MicroRNA. N Engl J Med. 2013;368(18):1685-1694. https://www.nejm.org/doi/full/10.1056/NEJMoa1209026 ↩︎ ↩︎ ↩︎ ↩︎
Foss F, et al. Phase 1 Trial of Cobomarsen (MRG-106), an Inhibitor of microRNA-155, in Patients with Cutaneous T-Cell Lymphoma. Blood. 2018;132(Suppl 1):2902. https://ashpublications.org/blood/article/132/supplement 1/2902/263360/phase-1-trial-of-cobomarsen-mrg-106-an-inhibitor ↩︎ ↩︎ ↩︎ ↩︎
Vermeire S, et al. ABX464 (Obefazimod) Upregulates miR-124 to Reduce Proinflammatory Markers in Inflammatory Bowel Diseases. Clin Transl Gastroenterol. 2023;14(4):e00572. https://pmc.ncbi.nlm.nih.gov/articles/PMC10132720/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Noureddine M, et al. Circulating microRNA profile of long-lived Okinawans identifies novel potential targets for optimizing lifespan and health span. Aging Cell. 2024;23(8):e14191. https://onlinelibrary.wiley.com/doi/10.1111/acel.14191 ↩︎ ↩︎
Yamakuchi M, Lowenstein CJ. MiR-34, SIRT1 and p53: the feedback loop. Cell Cycle. 2009;8(5):712-713. https://pubmed.ncbi.nlm.nih.gov/19221490/ ↩︎ ↩︎ ↩︎ ↩︎
Hatse S, et al. Circulating microRNAs as easy-to-measure aging biomarkers in older breast cancer patients: correlation with chronological age. PLoS One. 2014;9(10):e110644. https://journals.plos.org/plosone/article/file?type=printable&id=10.1371/journal.pone.0110644 ↩︎
Olivieri F, et al. Circulating inflamma-miRs in aging and age-related diseases. Front Genet. 2013;4:121. https://pmc.ncbi.nlm.nih.gov/articles/PMC3693036/ ↩︎
Dai M, et al. Safety and efficacy of the miR-124 upregulator ABX464 (obefazimod) in patients with active rheumatoid arthritis and inadequate response to methotrexate and/or anti-TNFα therapy: a placebo-controlled phase II study. Ann Rheum Dis. 2022;81(Suppl 1):321. https://pmc.ncbi.nlm.nih.gov/articles/PMC9279835/ ↩︎ ↩︎
Abplanalp WT, Fischer A, John D, et al. Efficiency and Target Derepression of Anti-miR-92a: Results of a First in Human Study. Nucleic Acid Therapeutics. 2020;30(6):335-342. https://journals.sagepub.com/doi/abs/10.1089/nat.2020.0871 ↩︎
Gallant-Behm CL, Piper J, Dickinson BA, et al. A synthetic microRNA-92a inhibitor (MRG-110) accelerates angiogenesis and wound healing in diabetic and nondiabetic wounds. Wound Repair Regen. 2018;26(4):311-323. https://pubmed.ncbi.nlm.nih.gov/30118158/ ↩︎
van Zandwijk N, Pavlakis N, Kao SC, et al. Safety and activity of microRNA-loaded minicells in patients with recurrent malignant pleural mesothelioma: a first-in-man, phase 1, open-label, dose-escalation study. Lancet Oncol. 2017;18(11):1456-1466. https://www.thelancet.com/journals/lanonc/article/PIIS1470-2045(17)30621-6/abstract ↩︎ ↩︎
Ipson BR, et al. Identifying exosome-derived microRNAs as candidate biomarkers of frailty. Sci Rep. 2018;8(1):15339. https://pmc.ncbi.nlm.nih.gov/articles/PMC6384524/ ↩︎ ↩︎ ↩︎ ↩︎
Zhao T, Li Z, Pang S, et al. P53/miR-34a/SIRT1 positive feedback loop regulates the termination of liver regeneration. Aging (Albany NY). 2021;13(4):5062-5079. https://www.aging-us.com/article/203920/text ↩︎ ↩︎
Zampetaki A, Kiechl S, Drozdov I, et al. Plasma MicroRNA profiling reveals loss of endothelial miR-126 and other microRNAs in type 2 diabetes. Circ Res. 2010;107(6):810-817. https://doi.org/10.1161/CIRCRESAHA.110.226357 ↩︎ ↩︎
de Lucia C, et al. MicroRNA-based therapeutics in cardiovascular disease. Eur J Intern Med. 2017;43:10-15. https://doi.org/10.1016/j.ejim.2017.05.011 ↩︎ ↩︎