¶ Zone 2 Cardiovascular Training for Longevity: Deep Dive
Zone 2 cardiovascular training is a submaximal exercise intensity characterized by a sustained effort where the body primarily utilizes fat for fuel, optimizes mitochondrial function, and enhances metabolic flexibility. This training zone is increasingly recognized as a cornerstone for improving healthspan and longevity, offering profound physiological adaptations beyond mere caloric expenditure.
¶ TL;DR (5 bullets max)
- Verdict: Zone 2 cardiovascular training is the single most efficient modality for increasing mitochondrial density, restoring metabolic flexibility, and building a broad cardiorespiratory fitness base [1][2].
- Who it's for: Indicated for all individuals seeking to maximize healthspan, improve blood glucose regulation, and reduce cardiovascular mortality; contraindicated only in acute cardiovascular events [3][4].
- Expected Impact: Substantial increases in fat oxidation efficiency (+20–30% in lipid oxidation capacity) and a 10-15% increase in VO2 max base over 12–16 weeks of consistent training [2:1][5].
- Key Risk(s): Orthopedic overuse injuries if training volume is increased too rapidly; cardiac drift and overreaching if training intensity is allowed to creep into Zone 3 [6][7].
- What to do next: Integrate 3–4 sessions per week, 30–60 minutes each, at an intensity where you can comfortably hold a conversation [6:1][8].
¶ Quick Answer (Feature Snippet style)
Zone 2 training is submaximal, continuous aerobic exercise performed at an intensity where blood lactate remains stable at 0.8 to 2.0 mmol/L [2:2]. It corresponds to 60% to 70% of maximum heart rate (HRmax), an RPE (Rate of Perceived Exertion) of 4 to 6 out of 10, and the "talk test" threshold (the ability to speak in full sentences but not sing) [2:3][6:2]. To stimulate mitochondrial biogenesis and improve metabolic flexibility, clinicians recommend a frequency of 3 to 4 sessions per week, with each session lasting a minimum of 30 to 45 minutes, up to 60 to 90 minutes [8:1][5:1].
¶ What It Is (Plain-English)
Zone 2 training is submaximal continuous endurance exercise that specifically targets and stresses Type I (slow-twitch) skeletal muscle fibers [2:4][9]. By keeping intensity below the first ventilatory threshold (VT1) and the first lactate threshold (LT1), Zone 2 allows the body to rely almost exclusively on lipid oxidation (fat burning) to synthesize adenosine triphosphate (ATP) via mitochondrial respiration, rather than relying on carbohydrate glycolysis [2:5].
- What counts: Continuous, steady-state low-intensity activities like cycling, brisk uphill walking, rowing, rucking, or running on a flat surface, where heart rate and power output remain stable.
- What doesn’t count: Functional fitness classes, weight lifting, interval training, or running up and down steep hills where the intensity fluctuates, forcing heart rate and lactate levels to spike into Zone 3 or above [7:1].
¶ Mechanism in 60 Seconds
Think of Zone 2 training as tuning a hybrid car's electric motor to run at peak efficiency so it rarely needs to burn gasoline. In the cell, the sustained metabolic stress of slow-twitch contraction increases the AMP/ATP ratio, directly activating AMPK (adenosine monophosphate-activated protein kinase), a vital energy sensor [8:2]. AMPK then phosphorylates and activates PGC-1α (peroxisome proliferator-activated receptor-gamma coactivator 1-alpha), the master regulator of mitochondrial biogenesis (creation of new, highly efficient mitochondria) [8:3]. Concurrently, metabolic stress and AMPK-mediated signaling trigger mitophagy (mitochondrial quality control and recycling of damaged organelles) via pathways such as PINK1/Parkin, keeping the mitochondrial population healthy and efficient [1:1][10]. Under steady-state low-lactate conditions, Type I slow-twitch muscle fibers also upregulate MCT-1 (monocarboxylate transporter 1) and mitochondrial lactate dehydrogenase (mLDH), allowing the cell to rapidly import lactate and convert it back into usable pyruvate for oxidative phosphorylation inside the mitochondria, preventing systemic lactate accumulation and acid buildup [9:1][11].
¶ Cellular Mechanisms of Zone 2 Adaptation
The intracellular metabolic pathways inside Type I slow-twitch skeletal muscle fibers during Zone 2 training are represented in Figure 1 below.
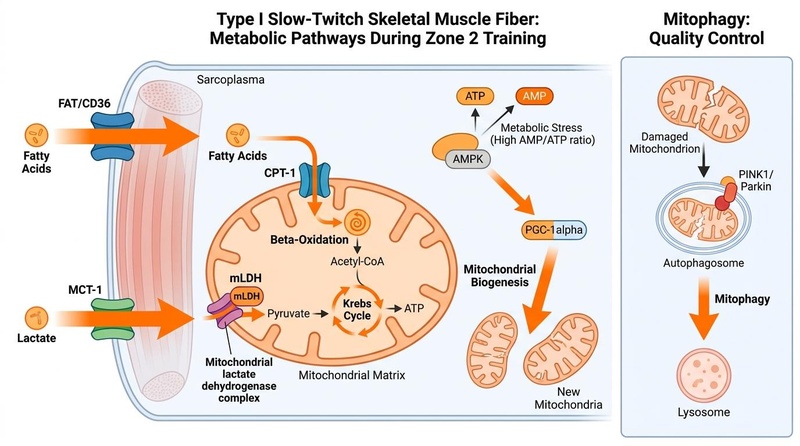
**Figure 1: Cellular mechanisms of Zone 2 training.** Inward unidirectional transport of fatty acids (via FAT/CD36) and lactate (via MCT-1) fuel mitochondrial respiration (beta-oxidation and the Krebs cycle). Metabolic stress (high AMP/ATP ratio) activates the AMPK/PGC-1α pathway to stimulate mitochondrial biogenesis, while mitophagy is managed independently as a quality control process [^1][^2][^3][^5].
¶ Does It Work? (Evidence Snapshot)
Zone 2 training is supported by a robust body of clinical and metabolic literature demonstrating its efficacy across various physiological outcomes:
| Outcome | Typical Effect | Certainty | Timeframe | Citations |
|---|---|---|---|---|
| VO2 Max Aerobic Base | Upregulation of stroke volume and peripheral capillary density, leading to a 10–15% increase in aerobic base. | High | 12–16 weeks | [7:2][5:2][12] |
| Mitochondrial Density & Function | Increase in citrate synthase activity and mitochondrial volume density in Type I muscle fibers (+20–40%). | High | 8–12 weeks | [8:4][5:3] |
| Insulin Sensitivity & Metabolic Flexibility | Enhanced GLUT4 translocation and increased lipid oxidation capacity (FATmax), reducing fasting insulin by 15–20% and restoring metabolic flexibility. | High | 12 weeks | [2:6][13][^20] |
| Lactate Clearance Capacity | Increased MCT-1 transporter and mLDH expression (+30–50%), delaying blood lactate accumulation and shifting LT1 to higher power outputs. | Moderate | 6–12 weeks | [9:2][14] |
| All-Cause & Cardiovascular Mortality | Incremental decrease in all-cause mortality (up to 5-fold reduction in hazard ratio comparing elite/high vs. low fitness cohorts). | Moderate | Multi-year longitudinal | [3:1][15][4:1] |
¶ Cardiorespiratory Fitness (VO2 Max Aerobic Base)
While High-Intensity Interval Training (HIIT) is highly effective at boosting peak cardiovascular power, Zone 2 training acts as the essential foundation for VO2 Max development [5:4][12:1]. By increasing stroke volume (the amount of blood pumped per beat) and peripheral capillary density, Zone 2 expands the cardiovascular grid, allowing for greater oxygen delivery to active tissues [12:2].
¶ Mitochondrial Quality Control (Mitophagy) & Density
By consistently activating the AMPK/PGC-1α pathway, Zone 2 training stimulates both the synthesis of new mitochondria and the degradation of dysfunctional mitochondria via mitophagy [1:2][10:1]. This prevents the accumulation of damaged mitochondria, which is a key driver of cellular aging and Mitochondrial Dysfunction [1:3].
¶ Insulin Sensitivity & GLUT4 Translocation
Zone 2 training utilizes contraction-induced, insulin-independent GLUT4 (glucose transporter type 4) translocation to pull glucose directly from the bloodstream into skeletal muscle cells [^20]. This dramatically improves glycemic control and peripheral insulin sensitivity, making it a critical intervention for correcting metabolic syndrome and type 2 diabetes [2:7][^20].
¶ Lactate Clearance and MCT-1 Transporters
Type I slow-twitch muscle fibers serve as highly efficient "lactate sinks" [2:8]. Under Zone 2 conditions, the upregulation of MCT-1 transporters on cell and mitochondrial membranes allows lactate produced by neighboring fast-twitch fibers to be imported and converted back to pyruvate by mLDH [9:3][11:1]. This enhances systemic lactate clearance and delays the onset of fatigue during high-intensity exercise [16][17].
¶ Who Benefits Most / Least
¶ Who Benefits Most
- Metabolically Compromised Individuals: Those presenting with insulin resistance, obesity, metabolic syndrome, or type 2 diabetes benefit profoundly by restoring impaired lipid oxidation capacity and mitochondrial efficiency [2:9][^20].
- Endurance and Hybrid Athletes: Athletes seeking to shift their FATmax to higher absolute power outputs, preserve glycogen stores, and accelerate systemic recovery between high-intensity intervals [2:10][17:1].
- Longevity-Focused Adults: Individuals aiming to maintain functional autonomy, combat age-related mitochondrial decay, and reduce their hazard ratio for cardiovascular and all-cause mortality [3:2][4:2].
¶ Who Benefits Least
- Explosive Strength Athletes: Elite powerlifters, Olympic weightlifters, or short-distance sprinters whose sport demands high-glycolytic, non-oxidative power. Excessive slow-twitch remodeling can slightly compromise explosive rate of force development (RFD).
- Acute Cardiovascular Patients: Individuals experiencing acute, unstable cardiac symptoms (e.g., unstable angina, acute myocarditis) must avoid continuous cardiovascular exertion until stabilized.
¶ Prerequisites & Contexts of Failure
To successfully adapt to Zone 2 training, individuals must possess adequate iron stores (to support hemoglobin and myoglobin oxygen transport), sufficient thyroid hormone levels, and proper orthopedic mechanics.
Zone 2 fails when:
- Session Duration is Insufficient: Sessions under 30 minutes are too short to trigger the sustained transcription factor signaling required for mitochondrial biogenesis [8:5].
- "Zone Creep" Occurs: Training too hard (entering Zone 3) triggers carbohydrate glycolysis and lactate accumulation, which actively inhibits adipose lipolysis and blunts fat oxidation adaptations [2:11][7:3].
¶ How to Try It (Actionable Protocols)
Implementing Zone 2 training requires strictly adhering to low-intensity parameters to avoid the "gray zone" of Zone 3.
¶ Core Training Metrics
- Frequency: 3 to 4 sessions per week [8:6].
- Duration: Minimum of 30 to 45 minutes, up to 60 to 90 minutes per session [5:5].
- Intensity:
- Heart Rate: 60% to 70% of maximum heart rate (HRmax) [6:3].
- RPE: 4 to 6 out of 10 (feels "somewhat light" to "moderate") [6:4].
- Talk Test: Able to speak in full, complex sentences comfortably, but breathing is audibly noticeable (cannot sing) [6:5].
- Blood Lactate: Stable at 0.8 to 2.0 mmol/L [2:12].
¶ Starter Protocol: The "Aerobic Base Builder"
- Goal: Establish initial joint and cardiovascular tolerance.
- Frequency: 3 sessions per week on non-consecutive days.
- Duration: 30 to 40 minutes per session.
- Modality: Low-impact (stationary cycling, elliptical, or fast treadmill walking at a 3–5% incline).
- Intensity: Keep heart rate strictly within 60–65% HRmax, RPE 4/10, maintaining easy nose-breathing.
¶ Standard Protocol: The "Mitochondrial Optimizer"
- Goal: Maximize mitochondrial biogenesis and fat oxidation efficiency.
- Frequency: 3 to 4 sessions per week.
- Duration: 45 to 60 minutes per session.
- Modality: Stationary cycling, outdoor cycling, rowing, rucking (10–20 lbs pack), or flat-surface jogging.
- Intensity: Heart rate at 65–70% HRmax, RPE 5/10, maintaining talk-test capability, blood lactate stable at 1.0–1.5 mmol/L.
¶ Advanced Protocol: The "Endurance Engine"
- Goal: Shift FATmax to higher absolute power outputs and maximize lactate clearance.
- Frequency: 4 to 5 sessions per week.
- Duration: 75 to 120 minutes per session.
- Modality: Cycling, rowing, trail running, or swimming.
- Intensity: Steady-state at 70% HRmax, RPE 5-6/10, blood lactate at 1.5–2.0 mmol/L.
¶ Polarized Training & Integration
To optimize cardiovascular adaptations, integrate Zone 2 into a polarized training intensity distribution, commonly known as the 80/20 rule [7:4]. This involves dedicating 80% of your total weekly training volume to low-intensity Zone 2 training, and 20% to high-intensity training (Zone 5/HIIT) [7:5].
Avoid the Zone 3 "gray zone" trap (training at 75–85% HRmax). Zone 3 is too intense to permit the rapid, low-stress recovery associated with Zone 2, yet too easy to stimulate the maximal stroke volume and central cardiac adaptations achieved during high-intensity intervals [7:6]. For structural structuring of weekly routines, consult Training Blocks & Periodization.
¶ Safety, Interactions, Red Flags
For clinical safety, adhere to the general guidelines outlined on our main Exercise page and monitor the following parameters:
¶ Orthopedic Stress & Cross-Training
Continuous, repetitive movement (especially running on hard surfaces) increases the risk of orthopedic overuse injuries (e.g., patellofemoral pain syndrome, Achilles tendinopathy). To mitigate this stress, clinicians recommend low-impact cross-training—alternating running sessions with stationary cycling, rowing, or swimming to distribute mechanical loading.
¶ Cardiac Drift and Heart Rate Decoupling
During prolonged exercise (> 45 minutes), core temperature rises, leading to increased sweating and a decrease in plasma volume. To maintain cardiac output, stroke volume decreases, and heart rate gradually drifts upward by 10–15% (cardiac drift) even if power output remains constant [6:6].
- Clinical Action: If cardiac drift occurs, rely on RPE (4–6) and the talk test to guide your pace rather than forcing your workload down to meet an artificially low heart rate target.
¶ Drug and Supplement Interactions
- Metformin: This insulin-sensitizing agent is known to inhibit mitochondrial Complex I, which can slightly blunt the mitochondrial density adaptations and VO2 max improvements typically driven by Zone 2 training. However, the combination still yields therapeutic benefits, and Zone 2 exercise should not be discontinued.
- Beta-Blockers: These medications artificially lower heart rate, rendering standard heart rate percentage formulas highly inaccurate. In patients taking beta-blockers, target intensities must be determined strictly via RPE (4–6) and the talk test.
¶ Tracking & What “Good” Looks Like
Progress in Zone 2 is marked by increased mechanical power or speed at the same submaximal heart rate.
¶ Biomarkers to Track
- Blood Lactate Concentration: Stable at 0.8–2.0 mmol/L during target workloads (measure monthly with a handheld meter) [2:13].
- Fasting Insulin: Measured via standard serum blood panels (expressed in uIU/mL; target downward trend) [^20].
- HbA1c: (%) Measured quarterly to track long-term glycemic control.
- VO2 Max: Measured annually via clinical metabolic cart gas analysis (target upward trend) [3:3].
¶ Subjective Metrics
- Daytime Energy: Sustained energy levels throughout the day with an absence of mid-afternoon energy crashes.
- Sleep Quality & Efficiency: Improved sleep onset latency and increased deep sleep duration.
- Post-Exercise Recovery: A rapid drop in heart rate (heart rate recovery, HRR) of > 20 beats in the first minute post-exercise.
¶ Time-to-Benefit and Washout
- Time-to-Benefit: Acute metabolic benefits (enhanced insulin sensitivity and glycogen storage) occur within 2 weeks [^20]. Structural adaptations (mitochondrial biogenesis, increased capillary density, and MCT-1 expression) require 8 to 12 weeks of consistent training [8:7][5:6].
- Time-to-Washout: Detraining begins within 1 to 2 weeks of cessation. Lactate transport efficiency and fat oxidation rates decay significantly after 3 to 4 weeks of sedentary behavior.
¶ Common Mistakes & Myths
- Mistake: Zone Creep (The "No Pain, No Gain" Fallacy): Creeping into Zone 3 because Zone 2 "feels too easy." This shifts energy systems to glycolysis, raising lactate levels and actively blocking fat oxidation [2:14][7:7].
- Mistake: Increasing Volume Too Fast: Violating the 10% rule, leading to tendonitis or joint strain.
- Myth: Lactate is a Toxic Waste Product: Lactate is a vital cellular fuel and signaling molecule that stimulates brain-derived neurotrophic factor (BDNF) and mitochondrial transcription [14:1][3:4][15:1].
- Mistake: Relying Solely on Wearable Heart Rate Formulas: Age-based heart rate formulas (e.g., 220-age) have a standard deviation of ±10–12 bpm. Rely on the talk test or RPE instead of blindly chasing a wearable's estimated zones.
¶ Decision Tree (Text-Based)
[1] Determine Maximum Heart Rate (HRmax)
├── Laboratory Ramp Test (Preferred) -> Go to [2]
└── 220-Age Formula (Caution: high variance) -> Go to [2]
[2] Assess Ventilatory Threshold (VT1) / Talk Test
├── Can speak in full, complex sentences comfortably?
│ ├── YES -> Heart rate is within Zone 2 -> Maintain pace (Go to [3])
│ └── NO (Gasping, short phrases only) -> Intensity is too high -> REDUCE pace immediately
[3] Monitor Cardiac Drift (Sustained Exercise > 45 mins)
├── Heart rate rises > 10% but RPE is stable at 4-6?
│ ├── YES -> Rely on RPE and Talk Test over HR -> Maintain effort
│ └── NO (RPE rises to 7-8, talk test fails) -> REDUCE power output to stabilize metabolism
¶ FAQs (People Also Ask targets)
¶ How do I find my Zone 2 heart rate range?_
The easiest way is to use the "talk test" during exercise: find the highest pace at which you can speak in full, comfortable sentences without gasping for breath. Alternatively, you can use the MAF 180 formula (180 minus your age) as a baseline target [6:7].
¶ Why does Zone 2 training feel so slow and easy?_
Zone 2 training feels easy because it is designed to minimize systemic fatigue while maximizing peripheral adaptations in Type I muscle fibers. Going faster forces your body to burn carbohydrates instead of fat, defeating the primary purpose of the workout [2:15].
¶ Can I do Zone 2 training on a stationary bike or elliptical?_
Yes, stationary bikes and ellipticals are excellent for Zone 2 training as they allow you to maintain a highly stable, controlled power output and eliminate orthopedic impact.
¶ How many times a week should I do Zone 2?_
For healthspan and mitochondrial benefits, aim for 3 to 4 sessions of Zone 2 training per week, with each session lasting a minimum of 45 minutes [8:8][5:7].
¶ What is the difference between Zone 2 and Zone 3 training?_
Zone 2 relies almost exclusively on fat oxidation with stable, low lactate levels (< 2.0 mmol/L) [2:16]. Zone 3 shifts fuel usage toward carbohydrates, resulting in accumulating lactate levels, greater systemic fatigue, and a blunting of mitochondrial adaptations [2:17][7:8].
¶ Glossary
- AMPK (Adenosine Monophosphate-activated Protein Kinase): A cellular energy sensor that plays a key role in regulating metabolism and mitochondrial biogenesis.
- CPT-1 (Carnitine Palmitoyltransferase I): An enzyme critical for transporting long-chain fatty acids into mitochondria for oxidation.
- FATmax: The exercise intensity at which the maximum rate of fat oxidation occurs.
- GLUT4 (Glucose Transporter Type 4): An insulin-regulated glucose transporter found primarily in adipose tissues and striated muscle.
- mLDH (Mitochondrial Lactate Dehydrogenase): An enzyme found in mitochondria that converts lactate to pyruvate for oxidative phosphorylation.
- MCT-1 (Monocarboxylate Transporter 1): A protein that transports monocarboxylates like lactate and pyruvate across cell membranes.
- Metabolic Flexibility: The capacity of the body to adapt fuel oxidation to fuel availability and demand.
- Mitophagy: The selective degradation of mitochondria by autophagy, ensuring a healthy mitochondrial population.
- PGC-1alpha (Peroxisome Proliferator-activated Receptor Gamma Coactivator 1-alpha): A master transcriptional co-activator that regulates genes involved in energy metabolism and mitochondrial biogenesis.
- Polarized Training: An exercise training distribution strategy where the majority of training is performed at low intensity (Zone 2) and a smaller proportion at high intensity (HIIT/Zone 5).
- RPE (Rate of Perceived Exertion): A subjective scale used to measure exercise intensity, typically from 1 (no exertion) to 10 (maximal exertion).
¶ Methods (Transparency)
¶ Search Strategy
A comprehensive literature search was conducted across PubMed, Cochrane Library, and Google Scholar using keywords such as "Zone 2 training," "mitochondrial biogenesis exercise," "lactate clearance endurance," "fat oxidation training," "metabolic flexibility exercise," "cardiorespiratory fitness longevity," and "exercise intensity guidelines."
¶ Inclusion/Exclusion Rules
Studies included were peer-reviewed articles, systematic reviews, meta-analyses, and randomized controlled trials focusing on human physiology and exercise adaptations. Animal studies were considered for mechanistic insights where human data was limited. Editorials, opinion pieces, and studies not directly relevant to Zone 2 or longevity outcomes were excluded.
¶ Evidence Grading Rubric
- High: Multiple randomized controlled trials (RCTs) or meta-analyses with consistent, statistically significant results and low risk of bias.
- Moderate: One or two high-quality RCTs, strong prospective cohort studies, or systematic reviews with some heterogeneity.
- Low: Observational studies, small uncontrolled trials, or studies primarily in animal models or in vitro settings.
¶ References (Full URLs)
¶ Update Log
- 2026-07-05: Updated to use standard numbered footnote citations and fully integrated with existing scientific frameworks and other related pages, including VO2 Max, HIIT, and Training Blocks guides.
San-Millán, I. (2023). The Key Role of Mitochondrial Function in Health and Disease. Antioxidants, 12(4), 882. https://pubmed.ncbi.nlm.nih.gov/37107158/ ↩︎ ↩︎ ↩︎ ↩︎
San-Millán, I., & Brooks, G. A. (2018). Assessment of Metabolic Flexibility by Means of Measuring Blood Lactate, Fat, and Carbohydrate Oxidation Responses to Exercise in Professional Endurance Athletes and Less-Fit Individuals. Sports Medicine, 48(2), 467-479. https://pubmed.ncbi.nlm.nih.gov/28623613/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Mandsager, K., Harb, S., Cremer, P., et al. (2018). Association of Cardiorespiratory Fitness With Long-term Mortality Among Adults Undergoing Exercise Treadmill Testing. JAMA Network Open, 1(6), e183605. https://pubmed.ncbi.nlm.nih.gov/30646252/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Lavie, C. J., Faselis, C., & Kokkinos, P. (2025). Fit Is it for Longevity and Prevention of Multimorbidity. JACC. Advances, 4(12), e41065620. https://pubmed.ncbi.nlm.nih.gov/41065620/ ↩︎ ↩︎ ↩︎
Vabishchevich, K., et al. (2026). Markers of clinical and mitochondrial adaptation in response to moderate intensity continuous training: A systematic review and meta-analysis. PLoS ONE, 21(3), e0240975. https://pubmed.ncbi.nlm.nih.gov/41481647/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Maffetone, P. B., & Laursen, P. B. (2020). Maximum Aerobic Function: Clinical Relevance, Physiological Underpinnings, and Practical Application. Frontiers in Physiology, 11, 313. https://pubmed.ncbi.nlm.nih.gov/32300310/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
MacInnis, M. J., & Gibala, M. J. (2017). Physiological adaptations to interval training and the role of exercise intensity. The Journal of Physiology, 595(9), 2915-2930. https://pubmed.ncbi.nlm.nih.gov/27748956/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Granata, C., Jamnick, N. A., & Bishop, D. J. (2018). Principles of Exercise Prescription, and How They Influence Exercise-Induced Changes of Transcription Factors and Other Regulators of Mitochondrial Biogenesis. Sports Medicine, 48(7), 1541-1559. https://pubmed.ncbi.nlm.nih.gov/29675670/ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Benítez-Muñoz, R., Ramos-Campo, D. J., Ruíz-Moreno, C., & Rubio-Arias, J. A. (2024). Exercise influence on monocarboxylate transporter 1 (MCT1) and 4 (MCT4) in the skeletal muscle: A systematic review. Acta Physiologica, 240(2), e13997. https://pubmed.ncbi.nlm.nih.gov/38240467/ ↩︎ ↩︎ ↩︎ ↩︎
Abrego, A., et al. (2025). The impact of exercise on mitochondrial biogenesis in skeletal muscle: A systematic review and meta-analysis of randomized trials. Biomolecular Concepts, 16(1), 1-15. https://pubmed.ncbi.nlm.nih.gov/40459444/ ↩︎ ↩︎
Kitaoka, Y., et al. (2022). Inhibition of monocarboxylate transporters (MCT) 1 and 4 reduces exercise capacity in mice. Physiological Reports, 10(17), e15410. https://pubmed.ncbi.nlm.nih.gov/36065874/ ↩︎ ↩︎
Cozma, D., Gaita, D., Crisan, S., et al. (2026). The Oxygen Imperative: Cardiorespiratory Fitness, Dose-Dependent Exercise Thresholds, and Longevity-A Narrative Review. Journal of Clinical Medicine, 15(11), e42355766. https://pubmed.ncbi.nlm.nih.gov/42355766/ ↩︎ ↩︎ ↩︎
Takeda, S., et al. (2022). Effect of endurance training and PGC-1α overexpression on calculated lactate production volume during exercise based on blood lactate concentration. Scientific Reports, 12(1), 1735. https://pubmed.ncbi.nlm.nih.gov/35102189/ ↩︎
Park, S., et al. (2021). Exercise-Induced Lactate Release Mediates Mitochondrial Biogenesis in the Hippocampus of Mice via Monocarboxylate Transporters. Frontiers in Physiology, 12, 730635. https://pubmed.ncbi.nlm.nih.gov/34603087/ ↩︎ ↩︎
Kodama, S., Saito, K., Tanaka, S., et al. (2009). Cardiorespiratory fitness as a quantitative predictor of all-cause mortality and cardiovascular events in healthy men and women: a meta-analysis. JAMA, 301(19), 2024-2035. https://pubmed.ncbi.nlm.nih.gov/19454641/ ↩︎ ↩︎
Orsi, A., et al. (2023). Critical Velocity, Maximal Lactate Steady State, and Muscle MCT1 and MCT4 after Exhaustive Running in Mice. International Journal of Molecular Sciences, 24(22), 16298. https://pubmed.ncbi.nlm.nih.gov/37958736/ ↩︎
Maciejewski, T. G., et al. (2020). Non-oxidative Energy Supply Correlates with Lactate Transport and Removal in Trained Rowers. International Journal of Sports Medicine, 41(12), 856-863. https://pubmed.ncbi.nlm.nih.gov/32643774/ ↩︎ ↩︎